Lecture notes on stability in the dynamics and Euler's method
 This semester, I had the opportunity to do 7 weeks of “interactive lectures” at QUT for MXB261 Modelling and Simulation Science, as well as one guest lecture on evolutionary game theory. In this blog post, I’ll talk about my experiences, share the first half of one of the lectures I gave, and provide the code, to generate the figures, in case any of that is useful to anyone. The Unit Coordinator is transforming the MXB261 into a game theory unit, and the first 7 weeks were “old” material she will replace in the coming years. Students responded really well to...
This semester, I had the opportunity to do 7 weeks of “interactive lectures” at QUT for MXB261 Modelling and Simulation Science, as well as one guest lecture on evolutionary game theory. In this blog post, I’ll talk about my experiences, share the first half of one of the lectures I gave, and provide the code, to generate the figures, in case any of that is useful to anyone. The Unit Coordinator is transforming the MXB261 into a game theory unit, and the first 7 weeks were “old” material she will replace in the coming years. Students responded really well to...Filed under: #teaching
Analytic solution for the stationary distribution in an iterated game with implementation errors
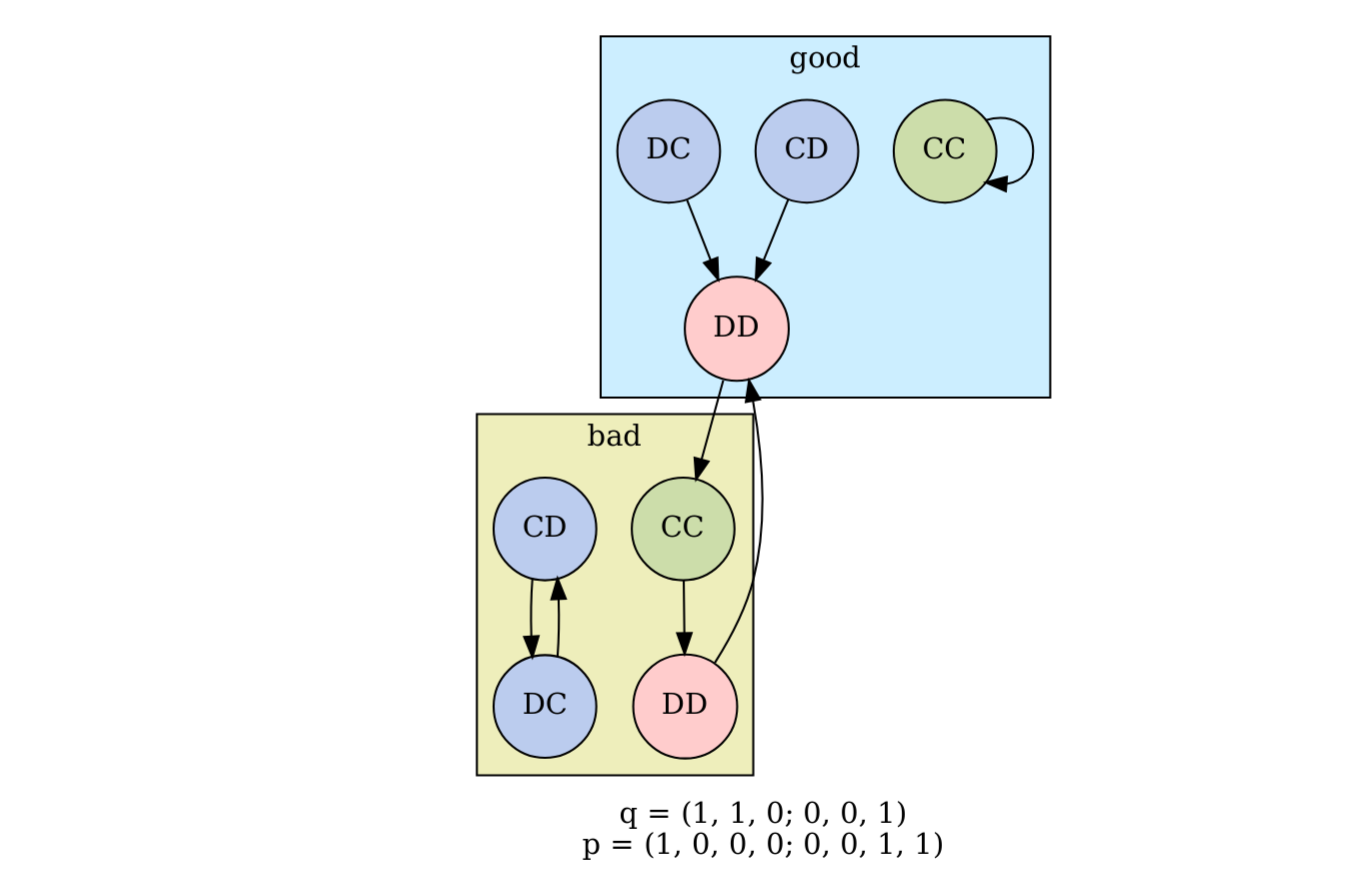 For the past year, I’ve been working in my spare time through a very interesting paper by Kleshnina et al. (2023). The paper concerns an evolutionary game theory model where individuals play an iterated Prisoner’s Dilemma with an environmental feedback. In a previous post, I experimented with a method to automate the identification of the subgame-perfect Nash equilibria and their parameter-value conditions. In this post, I’ll show how the stationary distribution of actions taken during a repeated interaction can be found analytically and solved algorithmically using SymPy. The typical approach requires choosing a specific value for \(\varepsilon\), which is the...
For the past year, I’ve been working in my spare time through a very interesting paper by Kleshnina et al. (2023). The paper concerns an evolutionary game theory model where individuals play an iterated Prisoner’s Dilemma with an environmental feedback. In a previous post, I experimented with a method to automate the identification of the subgame-perfect Nash equilibria and their parameter-value conditions. In this post, I’ll show how the stationary distribution of actions taken during a repeated interaction can be found analytically and solved algorithmically using SymPy. The typical approach requires choosing a specific value for \(\varepsilon\), which is the...Filed under: #cooperation
Evolution of cooperation talk at Griffith University
 Today,
I gave a talk at Griffith University’s
about my two most recent papers about the evolution of cooperation,
published in Scientific Reports
and Journal of Theoretical Biology.
You can download the full slide deck here: evoln_coopn.pdf.
Some slides from my talk
I have taken an adjunct position there, in addition to my position at
QUT,
and I’m looking forward to regular visits to the lovely Nathan Campus,
which is nestled in Toohey Forest.
Today,
I gave a talk at Griffith University’s
about my two most recent papers about the evolution of cooperation,
published in Scientific Reports
and Journal of Theoretical Biology.
You can download the full slide deck here: evoln_coopn.pdf.
Some slides from my talk
I have taken an adjunct position there, in addition to my position at
QUT,
and I’m looking forward to regular visits to the lovely Nathan Campus,
which is nestled in Toohey Forest.
Filed under: #cooperation
Cooperation among unequals
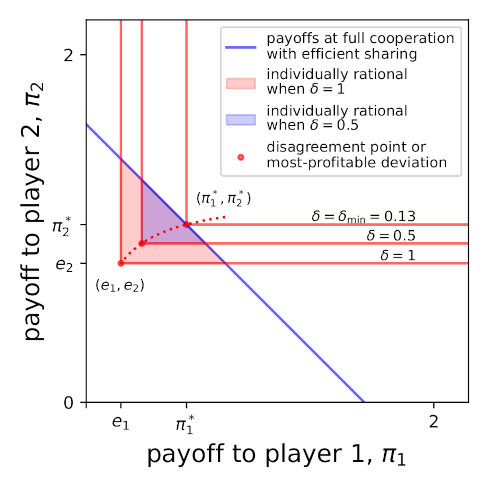 In an unpublished paper, Staab et al. (2022) studied the effects of heterogeneity in endowments and productivities on cooperation. They were particularly interested in the effect of allowing players to share the jointly produced goods unequally. Surprisingly, they found that unequal sharing can promote cooperation. In this blog post, first, I offer a proof of one of their propositions. Then I try thinking about their model a little differently to how they presented it, working through some small examples to try to get a visual intuition for what it means. Signs of eigenvalues proof Staab et al. (2022) model an...
In an unpublished paper, Staab et al. (2022) studied the effects of heterogeneity in endowments and productivities on cooperation. They were particularly interested in the effect of allowing players to share the jointly produced goods unequally. Surprisingly, they found that unequal sharing can promote cooperation. In this blog post, first, I offer a proof of one of their propositions. Then I try thinking about their model a little differently to how they presented it, working through some small examples to try to get a visual intuition for what it means. Signs of eigenvalues proof Staab et al. (2022) model an...Filed under: #cooperation
Evolutionary game theory virtual workshop
 This morning, I attended an engaging virtual workshop on modelling and applications of evolutionary game theory. I extend my sincere thanks to the organisers – Alex McAvoy, Olivia Chu, and Daniel Cooney (bsky) – for their kind invitation and organising the workshop. The presentations in my session were all very interesting. Maria Kleshnina's talk Maria Kleshnina presented an overview of her recent work on iterated games. I’m particularly intrigued by her model where player behaviour receives feedback from the environment, which I’ve previously discussed on my blog here. She also discussed the concept of endowment distributions and sharing rules that...
This morning, I attended an engaging virtual workshop on modelling and applications of evolutionary game theory. I extend my sincere thanks to the organisers – Alex McAvoy, Olivia Chu, and Daniel Cooney (bsky) – for their kind invitation and organising the workshop. The presentations in my session were all very interesting. Maria Kleshnina's talk Maria Kleshnina presented an overview of her recent work on iterated games. I’m particularly intrigued by her model where player behaviour receives feedback from the environment, which I’ve previously discussed on my blog here. She also discussed the concept of endowment distributions and sharing rules that...Filed under: #cooperation
Predicting the effects of species eradication from the community matrix alone
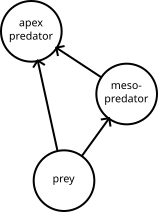 This blog post summarises one of the results I obtained from some work I did back in 2015 with Eve McDonald-Madden at the University of Queensland. She didn’t end up using it for anything, but perhaps someone else might find it useful. Modellers using the Qualitative Modelling approach often model a species eradication as a press-perturbation. This is obviously wrong, but it’s convenient and has a certain intuitive appeal, and it’s not known how often it gives answers that are wrong enough to matter. In this blog post, I provide a fast method to predict the effects of an eradication...
This blog post summarises one of the results I obtained from some work I did back in 2015 with Eve McDonald-Madden at the University of Queensland. She didn’t end up using it for anything, but perhaps someone else might find it useful. Modellers using the Qualitative Modelling approach often model a species eradication as a press-perturbation. This is obviously wrong, but it’s convenient and has a certain intuitive appeal, and it’s not known how often it gives answers that are wrong enough to matter. In this blog post, I provide a fast method to predict the effects of an eradication...Filed under: #qualitative_modelling
Check if an iterated Prisoner's Dilemma strategy is a subgame perfect Nash equilibrium
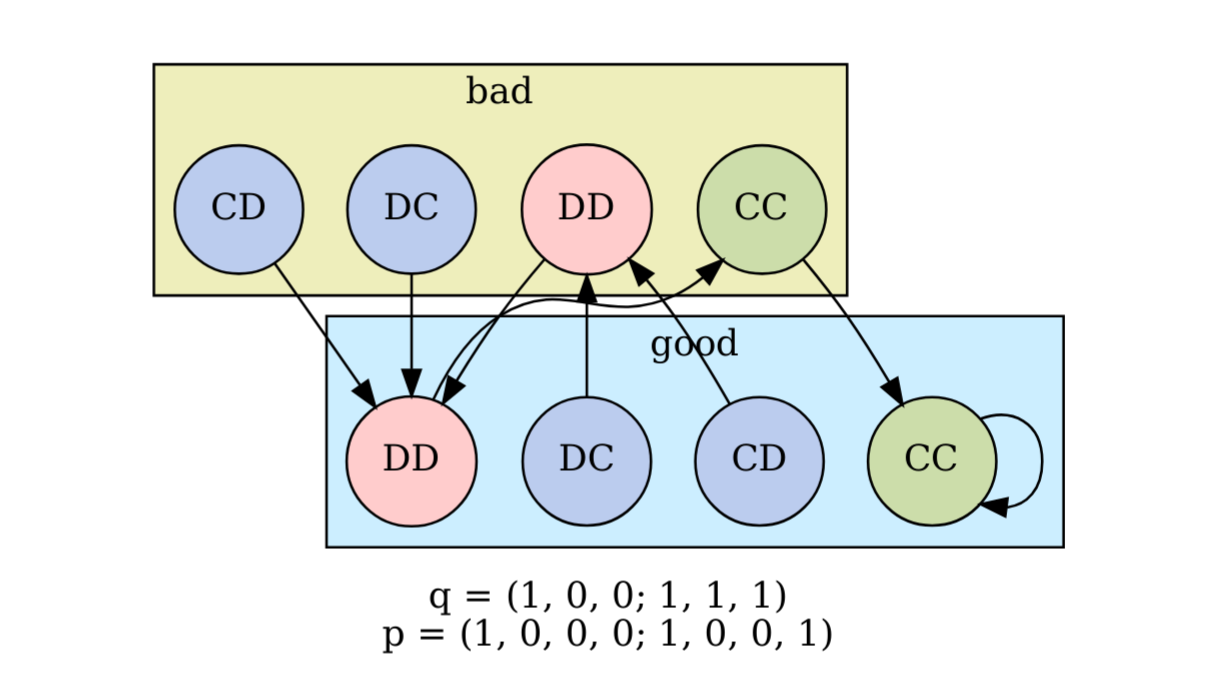 I recently read a paper by Kleshnina et al. (2023), The effect of environmental information on evolution of cooperation in stochastic games, which provided an opportunity to teach myself about how to analyse iterated games. In particular, the problem they investigated admits 64 possible scenarios with 256 possible strategies each, and I was interested in writing code that could automate the analysis. The solution I eventually landed on (Github repo) used a combination of SymPy, NetworkX, SageMath, the Z3 Theorem Prover, and PyEDA for Boolean minimisation, but I think my approach could be improved. Background to the paper Kleshnina et...
I recently read a paper by Kleshnina et al. (2023), The effect of environmental information on evolution of cooperation in stochastic games, which provided an opportunity to teach myself about how to analyse iterated games. In particular, the problem they investigated admits 64 possible scenarios with 256 possible strategies each, and I was interested in writing code that could automate the analysis. The solution I eventually landed on (Github repo) used a combination of SymPy, NetworkX, SageMath, the Z3 Theorem Prover, and PyEDA for Boolean minimisation, but I think my approach could be improved. Background to the paper Kleshnina et...Filed under: #cooperation
A summary of Richard Joyce's 'The Evolution of Morality'
 Metal. To learn more about the evolution of cooperation from a philosopher’s perspective, I recently read Richard Joyce’s book The Evolution of Morality. Joyce’s book makes the case for the evolutionary debunking argument, which holds that moral beliefs are the product of evolutionary processes rather than tracking moral truths. While evolution has equipped us with the capacity for moral judgement, this doesn’t necessarily mean that our moral beliefs are true or justified. Instead, our moral sense evolved because it was useful for our ancestors’ survival and reproduction, regardless of whether moral facts actually exist. Joyce begins by examining the evolutionary...
Metal. To learn more about the evolution of cooperation from a philosopher’s perspective, I recently read Richard Joyce’s book The Evolution of Morality. Joyce’s book makes the case for the evolutionary debunking argument, which holds that moral beliefs are the product of evolutionary processes rather than tracking moral truths. While evolution has equipped us with the capacity for moral judgement, this doesn’t necessarily mean that our moral beliefs are true or justified. Instead, our moral sense evolved because it was useful for our ancestors’ survival and reproduction, regardless of whether moral facts actually exist. Joyce begins by examining the evolutionary...Filed under: #cooperation
Coherence-based reasoning and the downplaying of contrary evidence
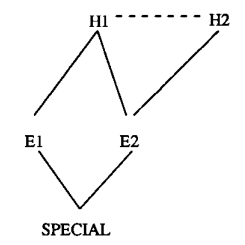 Human reasoning often defies the ideal of normative inference, where conclusions should logically follow from evidence. Instead, we frequently engage in ‘backward reasoning’, starting with preferred conclusions and selectively seeking supporting evidence while discounting contradictory information. This tendency reflects a broader cognitive preference for beliefs that align with our existing worldview. This inclination towards coherent beliefs contrasts sharply with foundationalist approaches to knowledge, which advocate building beliefs from self-evident truths (e.g., Descartes’ “I think; therefore, I am”). Our belief systems instead resemble intricate webs, characterised by complex interdependencies rather than clear hierarchies. Coherence-based reasoning is a cognitive model that attempts...
Human reasoning often defies the ideal of normative inference, where conclusions should logically follow from evidence. Instead, we frequently engage in ‘backward reasoning’, starting with preferred conclusions and selectively seeking supporting evidence while discounting contradictory information. This tendency reflects a broader cognitive preference for beliefs that align with our existing worldview. This inclination towards coherent beliefs contrasts sharply with foundationalist approaches to knowledge, which advocate building beliefs from self-evident truths (e.g., Descartes’ “I think; therefore, I am”). Our belief systems instead resemble intricate webs, characterised by complex interdependencies rather than clear hierarchies. Coherence-based reasoning is a cognitive model that attempts...Filed under: #cooperation
Guppy predator inspection
 I read a very interesting new paper by Padget et al. (2023), published late last year in Proceedings of the Royal Society B. The paper describes an experimental study of guppies’ predator-inspection behaviour, and it presents a bit of a mystery and a challenge to theoreticians. Many fishes perform predator-inspection, which is when a small number of individuals will break away from the main group and approach a potential predator (Dugatkin, 1988). Performing the inspection is risky behaviour, but it also benefits the group; it can provide the information needed to either escape or return foraging if there’s no real...
I read a very interesting new paper by Padget et al. (2023), published late last year in Proceedings of the Royal Society B. The paper describes an experimental study of guppies’ predator-inspection behaviour, and it presents a bit of a mystery and a challenge to theoreticians. Many fishes perform predator-inspection, which is when a small number of individuals will break away from the main group and approach a potential predator (Dugatkin, 1988). Performing the inspection is risky behaviour, but it also benefits the group; it can provide the information needed to either escape or return foraging if there’s no real...Filed under: #cooperation
Our new PNAS paper on historical extinctions in Singapore is out
 What are the impacts of tropical deforestation on biodiversity? To assess this, case studies are needed of tropical regions that have both a history of substantial deforestation and reliable biodiversity data. Singapore was almost entirely forested two hundred years ago but today less than 1% of the original primary forest remains today, with an additional ~20% of the landscape covered in lower quality secondary forest. Singapore also has an exceptionally detailed biodiversity record. Many species have gone extinct from Singapore, from the majestic tiger, the last of which was shot in the 1930s, to the humble epiphyte Hoya finlaysonii, which...
What are the impacts of tropical deforestation on biodiversity? To assess this, case studies are needed of tropical regions that have both a history of substantial deforestation and reliable biodiversity data. Singapore was almost entirely forested two hundred years ago but today less than 1% of the original primary forest remains today, with an additional ~20% of the landscape covered in lower quality secondary forest. Singapore also has an exceptionally detailed biodiversity record. Many species have gone extinct from Singapore, from the majestic tiger, the last of which was shot in the 1930s, to the humble epiphyte Hoya finlaysonii, which...Filed under: #undiscovered_extinctions
Web-based public goods game and other experiments for teaching
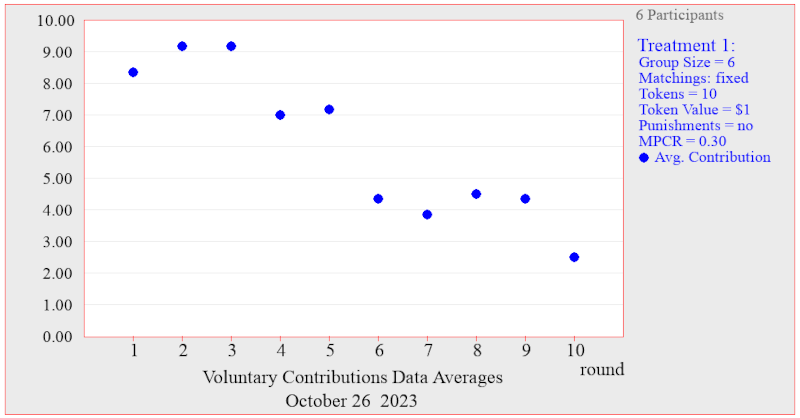 Each year since 2017, I’ve been taking one lecture for Ryan’s subject LSM4255 Methods in Mathematical Biology, teaching evolutionary game theory and adaptive dynamics. After the class, we usually have a tutorial where we play the fishing game, a simulation of a fishery that teaches students about the tragedy of the commons. Screenshot from the fishing game, which was sadly not available this year. Unfortunately, the fishing game was unexpectedly unavailable this year, but I was able to find a very nice last-minute alternative. Charles Holt at the University of Virginia has made available the Veconlab software, a collection of...
Each year since 2017, I’ve been taking one lecture for Ryan’s subject LSM4255 Methods in Mathematical Biology, teaching evolutionary game theory and adaptive dynamics. After the class, we usually have a tutorial where we play the fishing game, a simulation of a fishery that teaches students about the tragedy of the commons. Screenshot from the fishing game, which was sadly not available this year. Unfortunately, the fishing game was unexpectedly unavailable this year, but I was able to find a very nice last-minute alternative. Charles Holt at the University of Virginia has made available the Veconlab software, a collection of...Filed under: #cooperation #teaching
AMSI Winter School
 For 2 weeks, I’ve been teaching at and attending the AMSI Winter School at QUT (Mastodon thread about it here). The winter school is an annual event for postgrads, early-career researchers, and industry professionals, and about 40 students attended this year. Some students from the Winter School. I was one of four presenters on the theme Modelling Our Changing Biosphere. I taught about evolutionary game theory, and we also learnt about modelling fisheries (with Nokuthaba Sibanda from University of Wellington), reinforcement learning (with Carl Boettiger from University of California), model sloppiness (with Matthew Adams from Queensland University of Technology), and...
For 2 weeks, I’ve been teaching at and attending the AMSI Winter School at QUT (Mastodon thread about it here). The winter school is an annual event for postgrads, early-career researchers, and industry professionals, and about 40 students attended this year. Some students from the Winter School. I was one of four presenters on the theme Modelling Our Changing Biosphere. I taught about evolutionary game theory, and we also learnt about modelling fisheries (with Nokuthaba Sibanda from University of Wellington), reinforcement learning (with Carl Boettiger from University of California), model sloppiness (with Matthew Adams from Queensland University of Technology), and...Filed under: #teaching #cooperation
Visiting Hisashi Ohtsuki
 For the past 5 weeks, I’ve been visiting Hisashi Ohtsuki at SOKENDAI in Japan to work on our evolutionary game theory project. We’ve been collaborating with Hisashi over the past couple of years exploring questions related to the evolution of cooperation using a novel mathematical framework that accounts for higher-order genetic associations. This work resulted in a publication a few months ago. We’ve been putting the final touches on our extension of the higher-order genetic associations approach, to extend the scenario to games with many different strategies, and I’ll be here for another month so we can explore some new...
For the past 5 weeks, I’ve been visiting Hisashi Ohtsuki at SOKENDAI in Japan to work on our evolutionary game theory project. We’ve been collaborating with Hisashi over the past couple of years exploring questions related to the evolution of cooperation using a novel mathematical framework that accounts for higher-order genetic associations. This work resulted in a publication a few months ago. We’ve been putting the final touches on our extension of the higher-order genetic associations approach, to extend the scenario to games with many different strategies, and I’ll be here for another month so we can explore some new...Filed under: #cooperation
Evolution of cooperation
 When people play public goods games in lab-based experiments, they often cooperate even though it goes against their self interest. Many authors have pointed out the ways in which such experiments can be unrealistic, e.g., by enforcing anonymity between participants, and they have discussed how the incentives in a one-shot encounter differ from when interactions are repeated within the same group. In our recent paper, we focused on another common assumption that has received less attention: the benefits in real-life public goods games are almost never linear. We were interested in the implications of this nonlinearity for the evolution of...
When people play public goods games in lab-based experiments, they often cooperate even though it goes against their self interest. Many authors have pointed out the ways in which such experiments can be unrealistic, e.g., by enforcing anonymity between participants, and they have discussed how the incentives in a one-shot encounter differ from when interactions are repeated within the same group. In our recent paper, we focused on another common assumption that has received less attention: the benefits in real-life public goods games are almost never linear. We were interested in the implications of this nonlinearity for the evolution of...Filed under: #cooperation
Example numerical analysis of replicator dynamics with more than 2 strategies
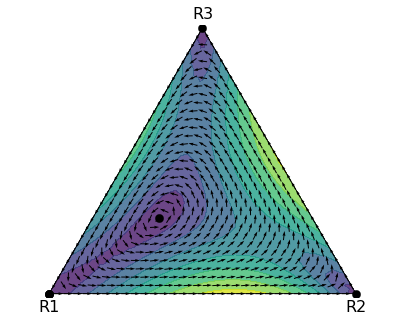 Recently, I set a task for a student to use Python to analyse the replicator dynamics of a game with three strategies, including using the Jacobian to determine the stability of the steady state. The purpose of this blog post is to share the solution in case that’s useful to someone. The game For this task, we will consider the replicator dynamics of a 2-player game with 3 strategies. Recall the general equation for the replicator dynamics is \[\begin{equation} \dot{p}_i = p_i (f_i - \bar{f}) \end{equation}\] where \(p_i\) is proportion of \(i\)-strategists in the population, \(f_i\) is fitness effect of...
Recently, I set a task for a student to use Python to analyse the replicator dynamics of a game with three strategies, including using the Jacobian to determine the stability of the steady state. The purpose of this blog post is to share the solution in case that’s useful to someone. The game For this task, we will consider the replicator dynamics of a 2-player game with 3 strategies. Recall the general equation for the replicator dynamics is \[\begin{equation} \dot{p}_i = p_i (f_i - \bar{f}) \end{equation}\] where \(p_i\) is proportion of \(i\)-strategists in the population, \(f_i\) is fitness effect of...Filed under: #cooperation #teaching
Group nepotism and the Brothers Karamazov Game
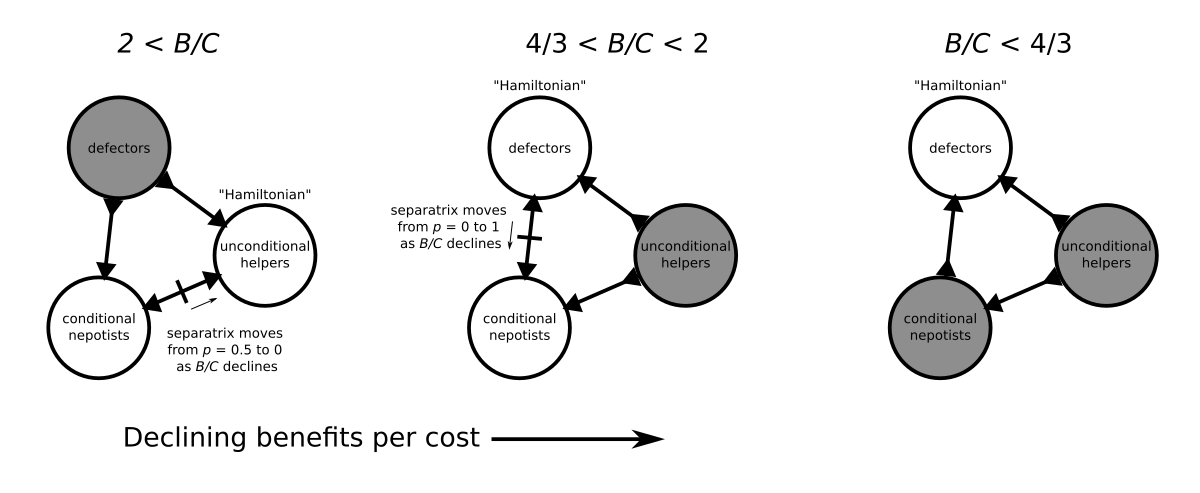 I recently read a classic paper by anthropologist Jones (2000), ‘Group nepotism and human kinship’, which argues that collective action may explain some of the key features of human cooperation between kin. Collective action here is action by a group (e.g., clan, tribe) that has sufficient ingroup solidarity to decide how to act as a collective. The key features it may help explain are: 1. the ‘axiom of amity’, that one is obliged to help kin just because they’re kin; 2. that kin groups come in many sizes and can be quite large; and 3. that human notions of kinship...
I recently read a classic paper by anthropologist Jones (2000), ‘Group nepotism and human kinship’, which argues that collective action may explain some of the key features of human cooperation between kin. Collective action here is action by a group (e.g., clan, tribe) that has sufficient ingroup solidarity to decide how to act as a collective. The key features it may help explain are: 1. the ‘axiom of amity’, that one is obliged to help kin just because they’re kin; 2. that kin groups come in many sizes and can be quite large; and 3. that human notions of kinship...Filed under: #cooperation
Lab meeting about threshold games
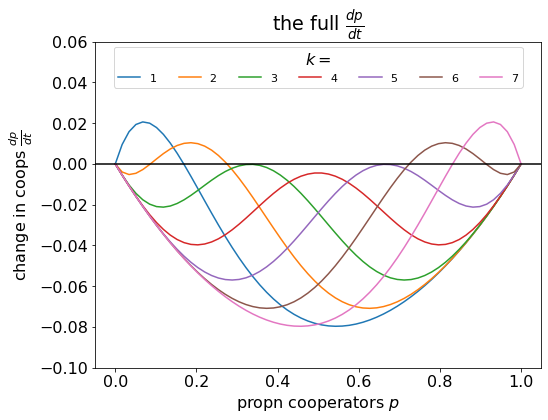 Last month, I chose a paper by Kris De Jaegher in Scientific Reports for our weekly lab-meeting discussion. We used the paper to teach ourselves about threshold games, and the purpose of this blog post is to summarise the things we learnt. Replicator dynamics revision De Jaegher used the replicator dynamics approach. Replicator dynamics assumes a well-mixed, infinitely large population of players that reproduces asexually, where the number of offspring they produce depends upon the payoff they receive from a game-theoretic ‘game’, as illustrated below:– Replicator dynamics animation, created by HowieKor (Creative Commons) We start with a population of \(n\)...
Last month, I chose a paper by Kris De Jaegher in Scientific Reports for our weekly lab-meeting discussion. We used the paper to teach ourselves about threshold games, and the purpose of this blog post is to summarise the things we learnt. Replicator dynamics revision De Jaegher used the replicator dynamics approach. Replicator dynamics assumes a well-mixed, infinitely large population of players that reproduces asexually, where the number of offspring they produce depends upon the payoff they receive from a game-theoretic ‘game’, as illustrated below:– Replicator dynamics animation, created by HowieKor (Creative Commons) We start with a population of \(n\)...Filed under: #cooperation #teaching
Predator dilution effect synchronises fish migration
 I recently came across a paper by Kaj Hulthen and others in Journal of Animal Ecology showing good empirical evidence for a model I coauthored a few years ago with Anna Harts and Hanna Kokko. Hulthen et al. (2022) were interested in the migration timing of roach (Rutilus rutilus). Specifically, they were interested in how the combination of selection for early arrival plus high predation risk could explain the relatively high migration synchrony in spring compared to autumn. Roach migrate from lake to stream in autumn, and from stream to lake in spring. In the lake in spring, zooplankton numbers...
I recently came across a paper by Kaj Hulthen and others in Journal of Animal Ecology showing good empirical evidence for a model I coauthored a few years ago with Anna Harts and Hanna Kokko. Hulthen et al. (2022) were interested in the migration timing of roach (Rutilus rutilus). Specifically, they were interested in how the combination of selection for early arrival plus high predation risk could explain the relatively high migration synchrony in spring compared to autumn. Roach migrate from lake to stream in autumn, and from stream to lake in spring. In the lake in spring, zooplankton numbers...Filed under: #evolutionary_ecology
Human cooperation and social network expansion over time
 It is generally held that human cooperation first evolved in our ancestral past, when we tended to live in small groups composed mostly of family members, and when cooperation could therefore be selected for by kin selection. This narrative is also sometimes invoked to explain the origin of cooperative mechanisms that are primarily about cooperation between nonkin. For example, in the iterated Prisoner’s Dilemma, cooperation between nonkin can be maintained by the tit-for-tat strategy, and a population of tit-for-tat players can resist invasion by defectors. However, tit-for-tat strategists cannot invade a population of defectors (presumably the primordial strategy), and so...
It is generally held that human cooperation first evolved in our ancestral past, when we tended to live in small groups composed mostly of family members, and when cooperation could therefore be selected for by kin selection. This narrative is also sometimes invoked to explain the origin of cooperative mechanisms that are primarily about cooperation between nonkin. For example, in the iterated Prisoner’s Dilemma, cooperation between nonkin can be maintained by the tit-for-tat strategy, and a population of tit-for-tat players can resist invasion by defectors. However, tit-for-tat strategists cannot invade a population of defectors (presumably the primordial strategy), and so...Filed under: #cooperation
Stochastic evolutionary dynamics of the Volunteers' Dilemma
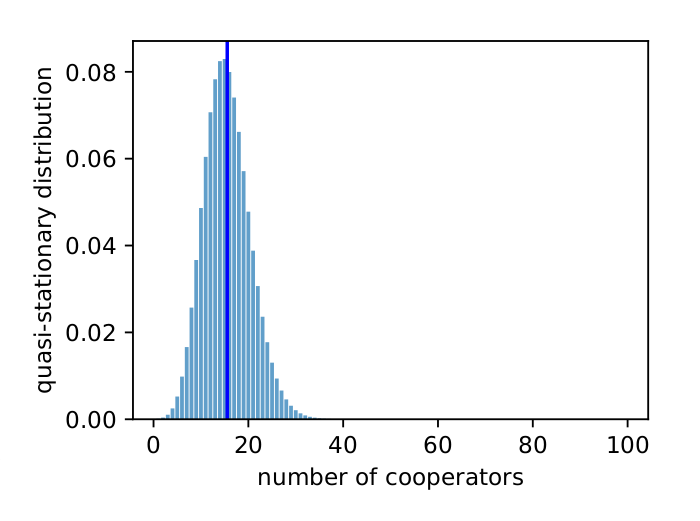 The purpose of this blog post is to use a recent paper by Tutić (2021) to teach myself about stochastic evolutionary game theory. Tutić’s (2021) model concerns the Volunteer’s Dilemma, which is a public goods game where the public good is provided if at least one group member cooperates. In the replicator dynamics, cooperators can always invade a population of defectors. However, as the group size increases, the proportion of cooperators in the population at the evolutionary steady-state declines (see Tutić (2021) Fig. 6), which increases the risk that cooperators will be lost from a finite population where stochastic forces...
The purpose of this blog post is to use a recent paper by Tutić (2021) to teach myself about stochastic evolutionary game theory. Tutić’s (2021) model concerns the Volunteer’s Dilemma, which is a public goods game where the public good is provided if at least one group member cooperates. In the replicator dynamics, cooperators can always invade a population of defectors. However, as the group size increases, the proportion of cooperators in the population at the evolutionary steady-state declines (see Tutić (2021) Fig. 6), which increases the risk that cooperators will be lost from a finite population where stochastic forces...Filed under: #cooperation
Using coalescence models to approximate interaction probabilities in weak selection
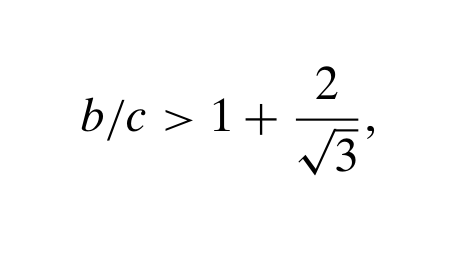 To calculate the conditions under which selection favours cooperation, Antal et al. (2009) used interaction probabilities calculated from a coalescence model. However, a coalescence model is a neutral model, it assumes no selective differences between types. So why can they assume no selective difference between types in a selective model? The purpose of this blog post is to explore the explanation detailed in their SI. First, a quick overview of the paper. The model involves the simultaneous evolution of two traits: (1) the strategy in a one-shot Prisoner’s Dilemma, either Cooperate or Defect; and (2) a phenotypic tag, modelled as...
To calculate the conditions under which selection favours cooperation, Antal et al. (2009) used interaction probabilities calculated from a coalescence model. However, a coalescence model is a neutral model, it assumes no selective differences between types. So why can they assume no selective difference between types in a selective model? The purpose of this blog post is to explore the explanation detailed in their SI. First, a quick overview of the paper. The model involves the simultaneous evolution of two traits: (1) the strategy in a one-shot Prisoner’s Dilemma, either Cooperate or Defect; and (2) a phenotypic tag, modelled as...Filed under: #cooperation
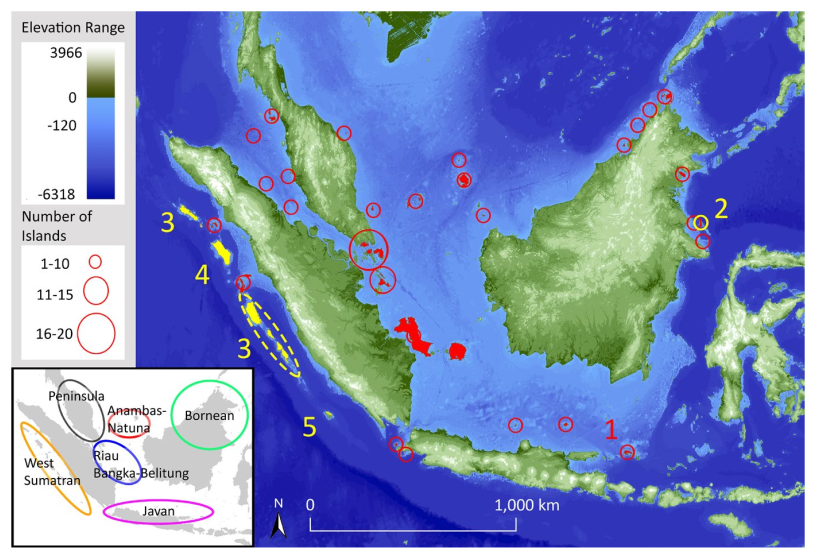
Island bird diversity not influenced by historical connection to the mainland
 The number of species on an island generally increases with island size and connectance to the mainland. However, if an island’s characteristics have changed in the recent past, that pattern will be disrupted. In Keita Sin’s recent paper in Journal of Biogeography, we tested if that expectation was borne out for birds on the Sundaic islands. Perhaps surprisingly, we found that it was not. The number of species on an island is generally determined by immigration and extinction (MacArthur & Wilson, 1967). Over long timescales, these two forces reach a stochastic equilibrium, producing the well-known species-richness patterns. Larger islands permit...
The number of species on an island generally increases with island size and connectance to the mainland. However, if an island’s characteristics have changed in the recent past, that pattern will be disrupted. In Keita Sin’s recent paper in Journal of Biogeography, we tested if that expectation was borne out for birds on the Sundaic islands. Perhaps surprisingly, we found that it was not. The number of species on an island is generally determined by immigration and extinction (MacArthur & Wilson, 1967). Over long timescales, these two forces reach a stochastic equilibrium, producing the well-known species-richness patterns. Larger islands permit...Filed under: #macroecology
Little difference between real food webs and randomly assembled webs
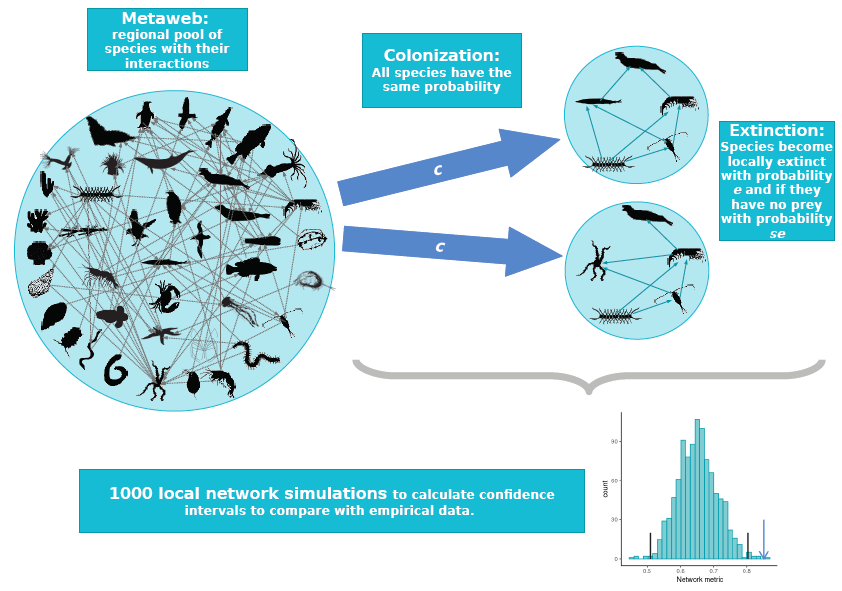 What determines the structure of a food web? It has long been theorised that food-web assembly is a selective process, allowing only certain species and interactions to persist in the face of dynamical stability constraints. In Leo Saravia’s recent paper in Journal of Animal Ecology, we used a novel test to see if we could detect the signal of this selection. We compared the structure of real food webs to those of webs randomly assembled from the regional metaweb. Contrary to theoretical expectations, we found almost no difference between them. Figure 1: The null food-web assembly model. Species randomly colonise...
What determines the structure of a food web? It has long been theorised that food-web assembly is a selective process, allowing only certain species and interactions to persist in the face of dynamical stability constraints. In Leo Saravia’s recent paper in Journal of Animal Ecology, we used a novel test to see if we could detect the signal of this selection. We compared the structure of real food webs to those of webs randomly assembled from the regional metaweb. Contrary to theoretical expectations, we found almost no difference between them. Figure 1: The null food-web assembly model. Species randomly colonise...Filed under: #food_webs
Connection between replicator dynamics, Bernstein polynomials, and Bezier curves
 A series of papers by Jorge Peña and others provide a neat way to characterise the replicator dynamics of two-strategy nonlinear public goods games with random group formation (Peña et al. 2014, Peña et al. 2015, Peña & Nöldeke 2016, Nöldeke & Peña 2016, Peña & Nöldeke 2018, Nöldeke & Peña 2020). I this blog post, I used a paper by Archetti (2018) to teach myself more about the topic. Archetti (2018) considered replicator dynamics in a population of producers (cooperators) and defectors. The probability that \(i\) of the \(n-1\) other members in your group are cooperators is \[p_{i,n-1}(x) =...
A series of papers by Jorge Peña and others provide a neat way to characterise the replicator dynamics of two-strategy nonlinear public goods games with random group formation (Peña et al. 2014, Peña et al. 2015, Peña & Nöldeke 2016, Nöldeke & Peña 2016, Peña & Nöldeke 2018, Nöldeke & Peña 2020). I this blog post, I used a paper by Archetti (2018) to teach myself more about the topic. Archetti (2018) considered replicator dynamics in a population of producers (cooperators) and defectors. The probability that \(i\) of the \(n-1\) other members in your group are cooperators is \[p_{i,n-1}(x) =...Filed under: #cooperation
Cooperation and maladaptation
Humans often cooperate in experimental games even when it’s not the strategy that will win them the most money (Ledyard, 1995; Cadsby and Maynes, 1999). They do this in one-shot games, where there is no chance the other could punish them if they took advantage, and in games with anonymous strangers, for whom they have no material reason to care. We see this cooperative tendency typically in the initial rounds of experimental public goods games (e.g., Offerman et al. 1996), but also in real-world common-resource problems (Ostrom 2000). From a naive economic or Darwinian perspective, which expects humans to behave...Filed under: #cooperation
Finding cell neighbours in an ISEA3H global grid in dggridR
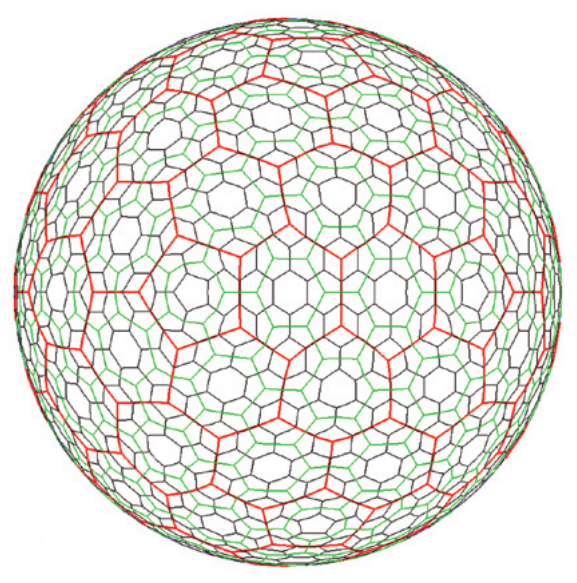 In order to model a spatially discretised process on the whole Earth, we need a discrete global grid system (DGGS). Ideally, the DGGS would divide the globe into a tessellation of some regular polygon, analogous to a grid of squares on the plane, where the grid resolution can be increased by further subdivision of the polygon. Another desirable property would be to have equal-distant centres, such as a hexagonal grid, so that the neighbourhood of a grid cell is well defined. Unfortunately, it is not possible to tessellate a sphere with hexagons only, and there are mathematical limits to e.g....
In order to model a spatially discretised process on the whole Earth, we need a discrete global grid system (DGGS). Ideally, the DGGS would divide the globe into a tessellation of some regular polygon, analogous to a grid of squares on the plane, where the grid resolution can be increased by further subdivision of the polygon. Another desirable property would be to have equal-distant centres, such as a hexagonal grid, so that the neighbourhood of a grid cell is well defined. Unfortunately, it is not possible to tessellate a sphere with hexagons only, and there are mathematical limits to e.g....Filed under: #coding #macroecology
Combining a stake in the future with the presence of others negatively affects intergenerational cooperation
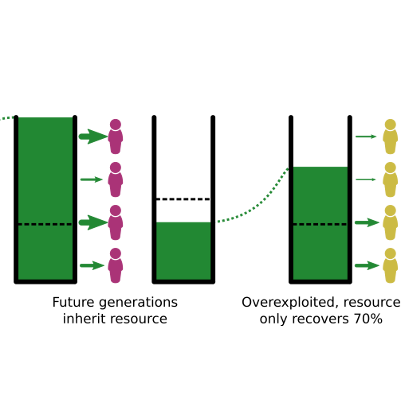 Conservationists often urge us to think about what kind of world we want to leave to future generations. But what if thinking about future generations — and about our descendants, specifically — triggers a geneological self-interest in us that can cause the environmental message to backfire? That was one of the questions raised by Chia-chen Chang’s new paper in Royal Society Open Science. The paper concerns results from a lab-based intergenerational common-pool resource game (Fischer et al. 2004, Hauser et al. 2014, Lohse et al. 2020). A common-pool resource game is a game that is designed to mimic the behaviour...
Conservationists often urge us to think about what kind of world we want to leave to future generations. But what if thinking about future generations — and about our descendants, specifically — triggers a geneological self-interest in us that can cause the environmental message to backfire? That was one of the questions raised by Chia-chen Chang’s new paper in Royal Society Open Science. The paper concerns results from a lab-based intergenerational common-pool resource game (Fischer et al. 2004, Hauser et al. 2014, Lohse et al. 2020). A common-pool resource game is a game that is designed to mimic the behaviour...Filed under: #cooperation
NODF nestedness worked example
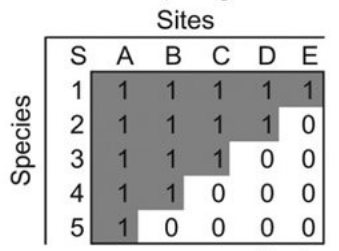 The purpose of this blog post is to work through the example in Almeida-Neto et al. (2008) for calculating the NODF nestedness metric. We’re interested in a species-site presence-absence matrix, which is a binary matrix with rows corresponding to species, columns corresponding to sites, and the presence of a species at a site represented by a 1. In a perfectly nested matrix, the sets of species at the sites and the sets of sites at which a species is present form proper subsets of each other. The figure below shows an example of a perfectly nested matrix. An example of...
The purpose of this blog post is to work through the example in Almeida-Neto et al. (2008) for calculating the NODF nestedness metric. We’re interested in a species-site presence-absence matrix, which is a binary matrix with rows corresponding to species, columns corresponding to sites, and the presence of a species at a site represented by a 1. In a perfectly nested matrix, the sets of species at the sites and the sets of sites at which a species is present form proper subsets of each other. The figure below shows an example of a perfectly nested matrix. An example of...Filed under: #macroecology
Intergenerational reciprocity and restoration
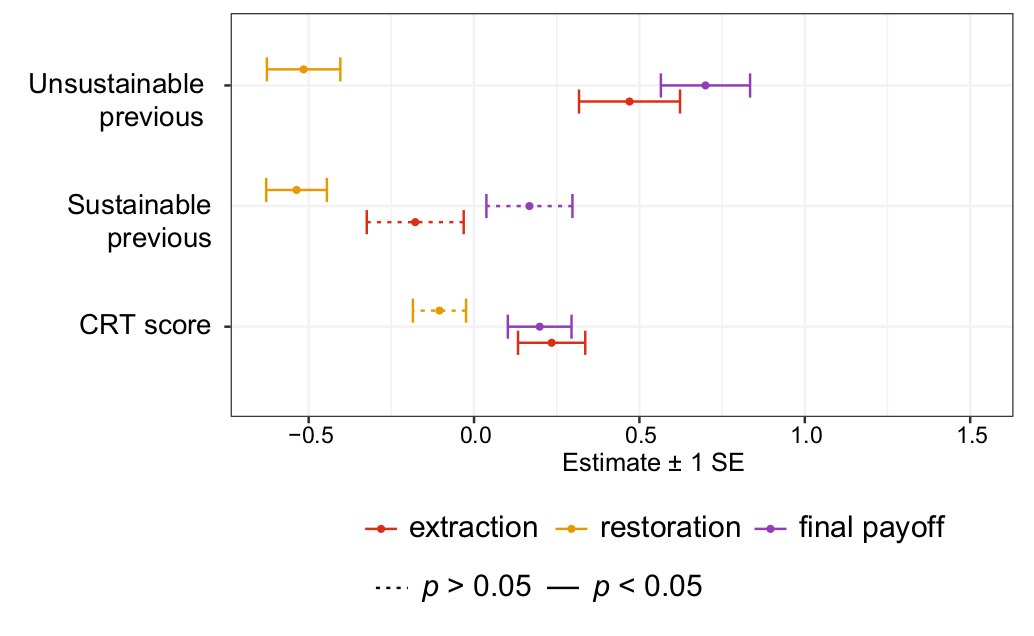 In conservation, we often talk about intergenerational equity, which is the idea that we should behave sustainably out of fairness towards future generations. However, how we treat others often depends on how we have been treated ourselves. If previous generations have over-exploited the natural resources and behaved unsustainably, will that make us feel less inclined to behave sustainably for future generations? In Chia-chen Chang’s recent paper in Sustainability Science, we explored this question using an experimental common-pool resource game. An example of a common-pool resource game The paper is specifically interested in restoration, and the key innovation was to investigate...
In conservation, we often talk about intergenerational equity, which is the idea that we should behave sustainably out of fairness towards future generations. However, how we treat others often depends on how we have been treated ourselves. If previous generations have over-exploited the natural resources and behaved unsustainably, will that make us feel less inclined to behave sustainably for future generations? In Chia-chen Chang’s recent paper in Sustainability Science, we explored this question using an experimental common-pool resource game. An example of a common-pool resource game The paper is specifically interested in restoration, and the key innovation was to investigate...Filed under: #cooperation
Homophily
Homophily is the principle that contact between similar people occurs at a higher rate than among dissimilar people (reviewed in McPherson et al., 2001). It is a long-established principle (see e.g., Byrne (1969), which has 1000+ citations), and is particularly well established for friendships, which have been shown to be homophilous for ethnicity, age, religion, education, and social values ([16,156–160] in Dunbar (2018)). ‘Baseline’ homophily occurs because of the demography of the potential relationship pool, and effects beyond that occur due to other factors not limited to personal preference. Homophily acts on a variety of different dimensions. The ‘Big 5’...Filed under: #cooperation
Evolutionary game theory model with homophilic imitation
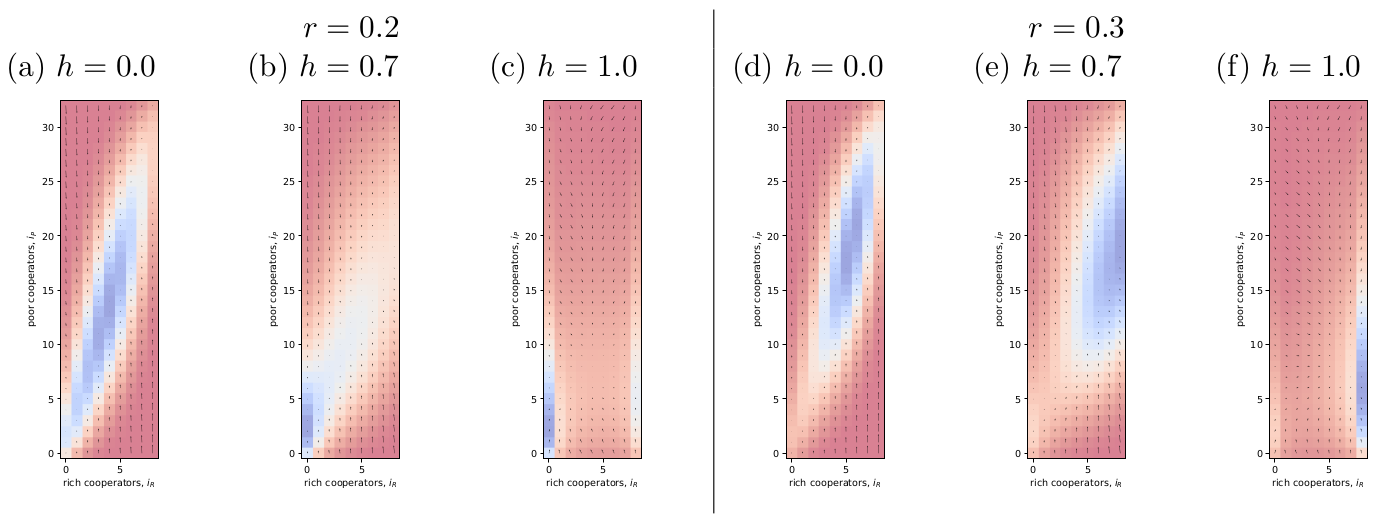 The purpose of this blog post is to replicate results from Vasconcelos et al. (2014) and use it to learn about the behaviour of this kind of model: Vasconcelos, V. V., Santos, F. C., Pacheco, J. M. and Levin, S. A. (2014). Climate policies under wealth inequality, Proceedings of the National Academy of Sciences 111(6): 2212–2216. I am interested in this model because it involves homophily: it is a social learning model, but the probability of imitation depends not only on the relative payoffs between the focal player and other but also on how similar they are in wealth. Code...
The purpose of this blog post is to replicate results from Vasconcelos et al. (2014) and use it to learn about the behaviour of this kind of model: Vasconcelos, V. V., Santos, F. C., Pacheco, J. M. and Levin, S. A. (2014). Climate policies under wealth inequality, Proceedings of the National Academy of Sciences 111(6): 2212–2216. I am interested in this model because it involves homophily: it is a social learning model, but the probability of imitation depends not only on the relative payoffs between the focal player and other but also on how similar they are in wealth. Code...Filed under: #cooperation
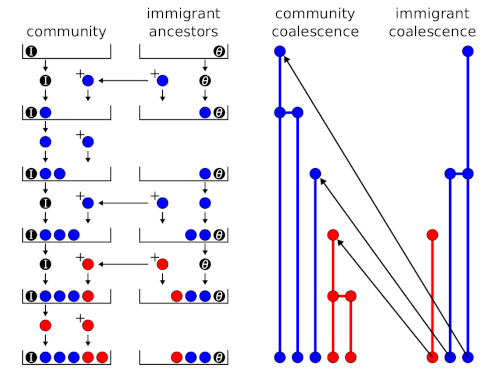
Sequential sampling schemes (urn models) for neutral ecological models
 A random sample of a species abundance distribution from a neutral metacommunity can be generated using a sequential sampling scheme or urn model (Etienne, 2005, 2007). An urn model is an idealised systems where some objects of actual statistical interest are represented as colored balls in an urn. In the ecological context, the balls represent individuals, the different colours are different species, and the urn’s final state represents the sample. I posted previously about the sequential sampling scheme that is used to generate a random sample from a neutral metacommunity. Hubbell (2001) called it a ‘species generator’, but the concept...
A random sample of a species abundance distribution from a neutral metacommunity can be generated using a sequential sampling scheme or urn model (Etienne, 2005, 2007). An urn model is an idealised systems where some objects of actual statistical interest are represented as colored balls in an urn. In the ecological context, the balls represent individuals, the different colours are different species, and the urn’s final state represents the sample. I posted previously about the sequential sampling scheme that is used to generate a random sample from a neutral metacommunity. Hubbell (2001) called it a ‘species generator’, but the concept...Filed under: #macroecology
Using n-player relatedness to model the Modern Tragedy of the Commons
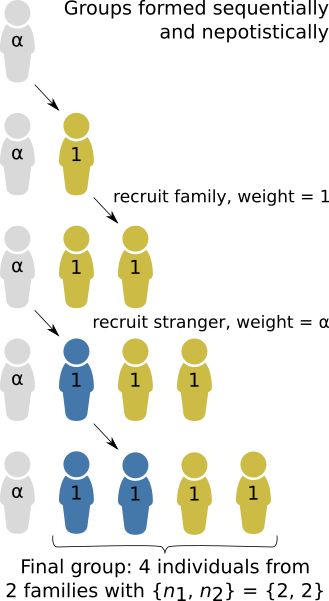 In this blog post, I will use the “Modern Tragedy of the Commons” as an example for how to use Hisashi Ohtsuki’s \(n\)-player generalisation of relatedness. In a recent experimental study in Science Advances, Gross and De Dreu (2019) described the Modern Tragedy of the Commons: if people can individually solve a problem by purchasing a solution that protects themselves only, then they may prefer that to contributing to a cheaper collective solution (Fig. 1). One recent example might be the trend among wealthy Californian home-owners to hire private firefighters to defend their property from wildfires exacerbated by climate change....
In this blog post, I will use the “Modern Tragedy of the Commons” as an example for how to use Hisashi Ohtsuki’s \(n\)-player generalisation of relatedness. In a recent experimental study in Science Advances, Gross and De Dreu (2019) described the Modern Tragedy of the Commons: if people can individually solve a problem by purchasing a solution that protects themselves only, then they may prefer that to contributing to a cheaper collective solution (Fig. 1). One recent example might be the trend among wealthy Californian home-owners to hire private firefighters to defend their property from wildfires exacerbated by climate change....Filed under: #cooperation
Where does Hubbell's 'species generator' come from?
On page 289 onwards, Hubbell (2001) gives an algorithm for sequentially sampling individuals from the neutral metacommunity and obtaining their species identities. The algorithm makes use of a quantity he calls the ‘species generator’ \[\frac{\theta}{\theta + j - 1}\] where \(\theta\) is the “fundamental biodiversity number” and \(j\) is the index of the individual drawn (1st, 2nd, …). In Figure 9.1 he gives a flow diagram for the algorithm, but it is perhaps easier to understand from reading code directly. From the hubbell package for R, Jari Oksanen authored this code: function(theta, J) { community <- NULL for (j in...Filed under: #macroecology
Estimating undiscovered extinctions
 Have you ever wondered how many species were lost before we had the chance to discover them? In a paper now out in Conservation Biology, we estimated just that, for plant species in Singapore. This paper follows from the Chisholm Lab’s related work on Singapore birds and butterflies. The orchid species Grammatophyllum speciosum has not been recorded in Singapore since 1918. Photo credit: Cerlin Ng All over the world, many species remain undiscovered while both known and unknown species continue to go extinct. This is particularly true in the tropics, where biodiversity is high and development continues apace. Singapore provides...
Have you ever wondered how many species were lost before we had the chance to discover them? In a paper now out in Conservation Biology, we estimated just that, for plant species in Singapore. This paper follows from the Chisholm Lab’s related work on Singapore birds and butterflies. The orchid species Grammatophyllum speciosum has not been recorded in Singapore since 1918. Photo credit: Cerlin Ng All over the world, many species remain undiscovered while both known and unknown species continue to go extinct. This is particularly true in the tropics, where biodiversity is high and development continues apace. Singapore provides...Filed under: #undiscovered_extinctions
Boolean qualitative modelling on Christmas Island
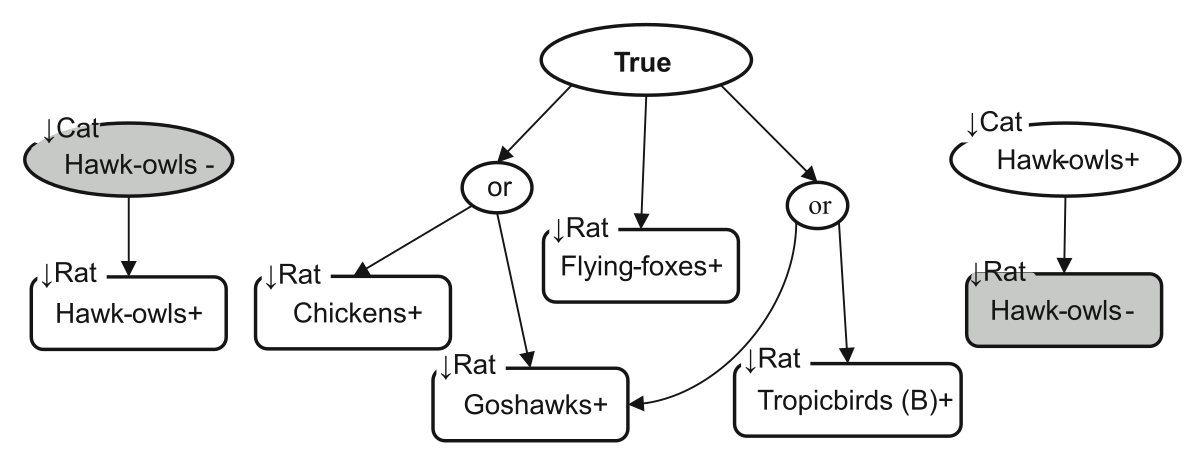 Conservation managers often want us to predict how species will respond to different management options when we don’t have enough data to parameterise a dynamical model. Previously, we found that a popular Qualitative Modelling technique involving probabilistic analysis has philosophical problems and produces contradictory results. We proposed a new method, based on Boolean analysis, that remedied these problems (Kristensen et al. 2019 Meth Ecol Evol). However, it was still an open question how the Boolean approach would perform in a real-world application. Yi Han’s new paper in Ecological Modelling is the first time the Boolean approach has been applied to...
Conservation managers often want us to predict how species will respond to different management options when we don’t have enough data to parameterise a dynamical model. Previously, we found that a popular Qualitative Modelling technique involving probabilistic analysis has philosophical problems and produces contradictory results. We proposed a new method, based on Boolean analysis, that remedied these problems (Kristensen et al. 2019 Meth Ecol Evol). However, it was still an open question how the Boolean approach would perform in a real-world application. Yi Han’s new paper in Ecological Modelling is the first time the Boolean approach has been applied to...Filed under: #qualitative_modelling
Extended Tea Bag Index to measure microbial- and termite-driven decomposition in the tropics
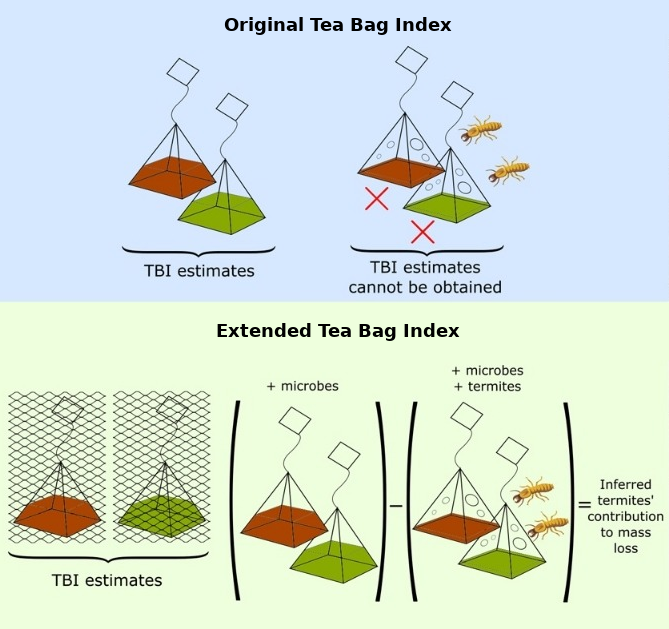 Our new paper about using the Tea Bag Index in the tropics, headed by Aloysius Teo, has just come out in Pedobiologia. The Tea Bag Index (TBI) was designed by Keuskamp et al. (2013) as a standardised and cheap method to quantify microbial-driven decomposition by measuring the mass loss of tea within tea bags. It can be used by scientists and citizen-scientists alike. School students from cpt. Nálepka elementary school, Slovakia, participate in Teatime4science. (Photo credit) However, we found that when the TBI was deployed in tropical forests, termites damaged up to 80% of the tea bags. Termites also consumed...
Our new paper about using the Tea Bag Index in the tropics, headed by Aloysius Teo, has just come out in Pedobiologia. The Tea Bag Index (TBI) was designed by Keuskamp et al. (2013) as a standardised and cheap method to quantify microbial-driven decomposition by measuring the mass loss of tea within tea bags. It can be used by scientists and citizen-scientists alike. School students from cpt. Nálepka elementary school, Slovakia, participate in Teatime4science. (Photo credit) However, we found that when the TBI was deployed in tropical forests, termites damaged up to 80% of the tea bags. Termites also consumed...Filed under: #other
Example debugging mixed Python C++ in VS Code
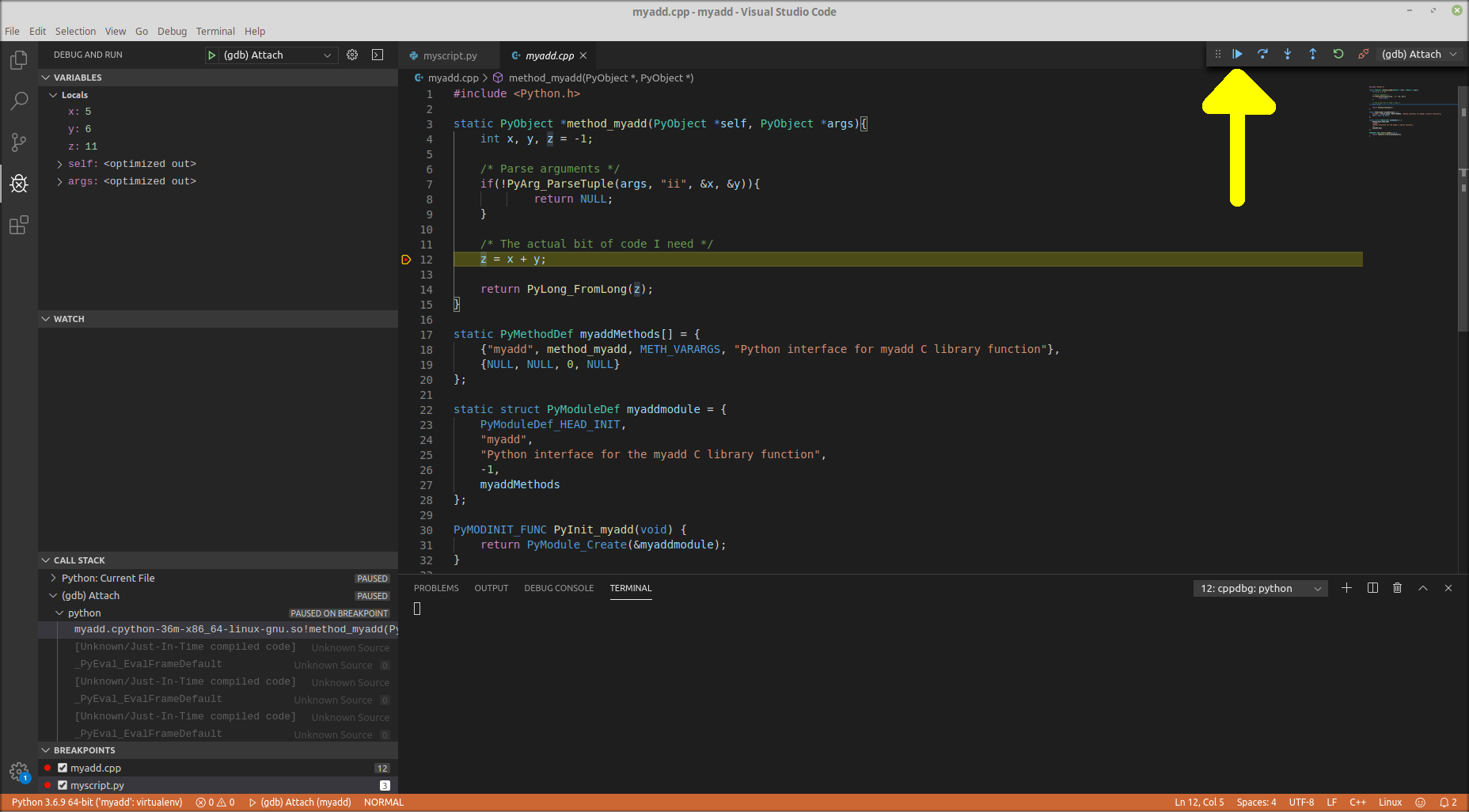 Visual Studio Code has the ability to debug mixed Python with C++ extensions. In this blog post, I give an example of how to get it working. I’m going to do the example from scratch in five steps: Make virtual environment. Chances are that, if you’re doing this kind of thing, you’ll be wanting to use a virtual environment too. Write code. My toy example is a C++ extension that just adds two numbers together. Set up VS Code project. Important to point the interpreter to virtual environment. Create launch.json. Configure the debugger so it can both run on python...
Visual Studio Code has the ability to debug mixed Python with C++ extensions. In this blog post, I give an example of how to get it working. I’m going to do the example from scratch in five steps: Make virtual environment. Chances are that, if you’re doing this kind of thing, you’ll be wanting to use a virtual environment too. Write code. My toy example is a C++ extension that just adds two numbers together. Set up VS Code project. Important to point the interpreter to virtual environment. Create launch.json. Configure the debugger so it can both run on python...Filed under: #coding
Transient dynamics in neutral models
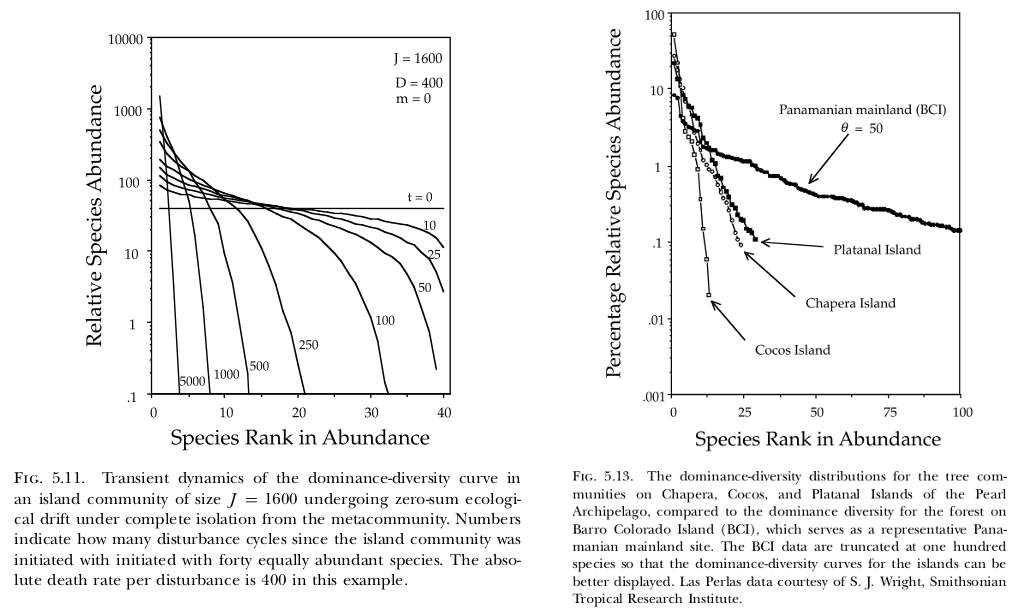 In their 10-year-anniversary review of neutral theory (Hubbell, 2001), Rosindell et al. (2011) note that little work has been done on the model’s transient dynamics. Transient dynamics are nonetheless interesting because they can capture important processes occurring in recently disturbed landscapes. For example, if forest is cleared such that only a small patch remains, then the number of species in that patch will continue to decline over time (i.e. extinction debt). As another example (Hubbell, 2001, p. 140–142), the Pearl Archipelago in the Bay of Panama was once a continuous coastal plain attached to the mainland. As the water rose...
In their 10-year-anniversary review of neutral theory (Hubbell, 2001), Rosindell et al. (2011) note that little work has been done on the model’s transient dynamics. Transient dynamics are nonetheless interesting because they can capture important processes occurring in recently disturbed landscapes. For example, if forest is cleared such that only a small patch remains, then the number of species in that patch will continue to decline over time (i.e. extinction debt). As another example (Hubbell, 2001, p. 140–142), the Pearl Archipelago in the Bay of Panama was once a continuous coastal plain attached to the mainland. As the water rose...Filed under: #macroecology
Extinction of undiscovered butterflies + tutorial
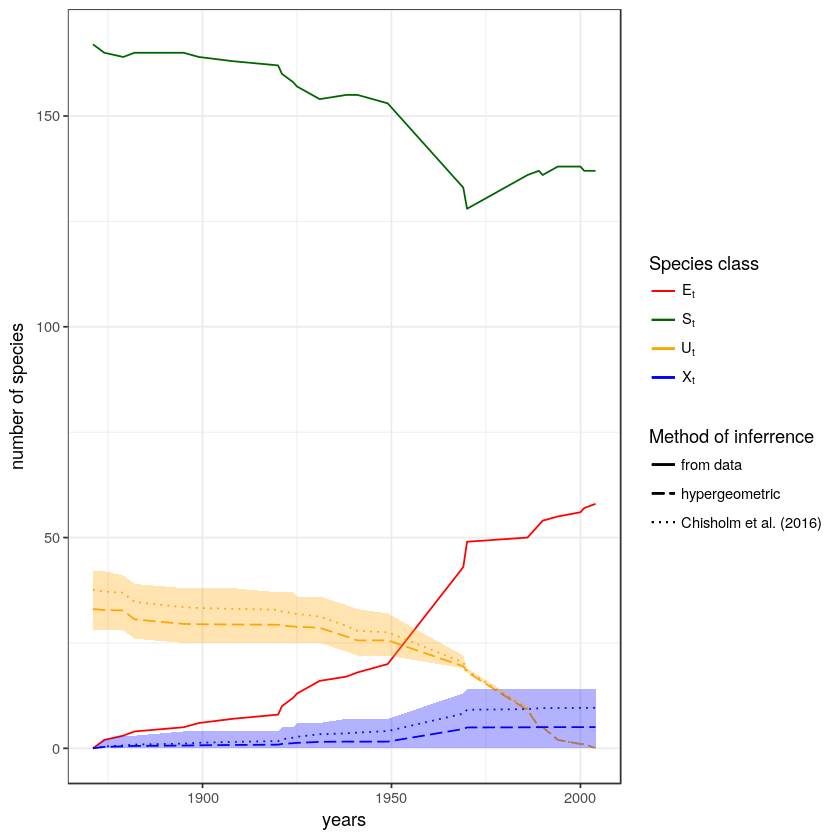 Meryl Theng just had a new paper published in Biological Conservation, where she estimated that 46% of Singapore’s butterfly species have been extirpated since 1854. The special thing about this estimate is that it includes all species that existed, including species that went extinct before we had a chance to discover them. The trick to estimating undiscovered extinctions is the SEUX model. There is a nice write-up about the paper on Ryan’s blog. The paper has also received a good response in the press - The Straits Times and The Star have covered it - and it is generating some...
Meryl Theng just had a new paper published in Biological Conservation, where she estimated that 46% of Singapore’s butterfly species have been extirpated since 1854. The special thing about this estimate is that it includes all species that existed, including species that went extinct before we had a chance to discover them. The trick to estimating undiscovered extinctions is the SEUX model. There is a nice write-up about the paper on Ryan’s blog. The paper has also received a good response in the press - The Straits Times and The Star have covered it - and it is generating some...Filed under: #coding #undiscovered_extinctions
New solutions for Parker sperm competition model
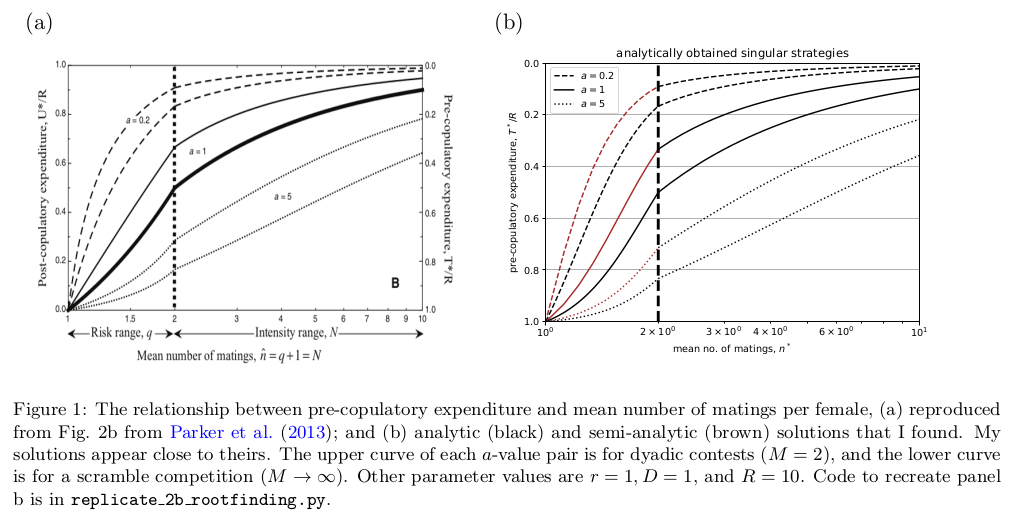 Parker et al. (2013) created a general model for sperm allocation under a trade-off between male investment of resources \(R\) into pre-copulatory effort (e.g. search time) \(T\) versus post-copulatory effort (e.g. ejaculate) \(U\). Their model is interesting because it encompasses a range of different scenarios of female remating and the type of competition between males. For female remating scenarios, the risk model has females mating a second time with probability \(q\), with the possibility that being first or second leads to higher fertilisation rate (`loaded raffles’); whereas the intensity model has females mating with \(N > 2\) males on average....
Parker et al. (2013) created a general model for sperm allocation under a trade-off between male investment of resources \(R\) into pre-copulatory effort (e.g. search time) \(T\) versus post-copulatory effort (e.g. ejaculate) \(U\). Their model is interesting because it encompasses a range of different scenarios of female remating and the type of competition between males. For female remating scenarios, the risk model has females mating a second time with probability \(q\), with the possibility that being first or second leads to higher fertilisation rate (`loaded raffles’); whereas the intensity model has females mating with \(N > 2\) males on average....Filed under: #other
Playing with a new model for fugitive coexistence
 I recently read a paper by Kawecki (2017), which presents a new mechanism for something analogous to fugitive coexistence. The paper has a really great literature overview, which I won’t be able to do justice here. In short, fugitive coexistence is when an inferior species persists on a patchy landscape by being a better coloniser: when a local extinction occurs, they are quicker to arrive at the patch and so exploit the time window before the superior competitor arrives. But Kawecki had another idea about how coexistence could occur. In the paper, Kawecki explores the coexistence of a ‘jack of...
I recently read a paper by Kawecki (2017), which presents a new mechanism for something analogous to fugitive coexistence. The paper has a really great literature overview, which I won’t be able to do justice here. In short, fugitive coexistence is when an inferior species persists on a patchy landscape by being a better coloniser: when a local extinction occurs, they are quicker to arrive at the patch and so exploit the time window before the superior competitor arrives. But Kawecki had another idea about how coexistence could occur. In the paper, Kawecki explores the coexistence of a ‘jack of...Filed under: #evolutionary_ecology #dispersal
Typo in qualitative modelling paper
There is a typo in our recent paper to MEE, Dealing with high uncertainty in qualitative network models using Boolean analysis. Example 2 of Box 1 should read “y = water/wine” not “y = wine/water”. Many thanks to Anubhav Gupta at University of Zurich for emailing us to let us know.Filed under: #qualitative_modelling
Boolean approach to qualitative network modelling
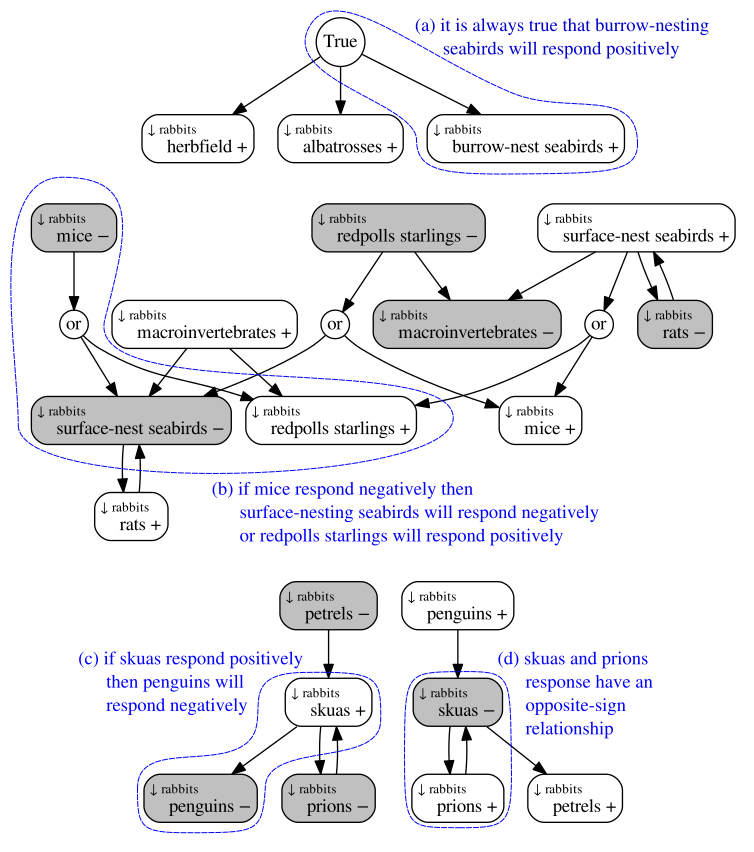 In a new paper in Methods in Ecology and Evolution (draft with supplementary here), we tackled an important question for ecological modellers: how do we predict an ecosystem’s behaviour when the data needed to parameterise a model are lacking? This problem is particularly important when our models are needed for conservation decision-making. For example, managers may be considering different pest-control programmes, which have the potential to lead to negative outcomes for native species, and predictions of these outcomes are needed for sound decisions to be made. These outcomes can in principle be predicted using dynamical models, however experts rarely have...
In a new paper in Methods in Ecology and Evolution (draft with supplementary here), we tackled an important question for ecological modellers: how do we predict an ecosystem’s behaviour when the data needed to parameterise a model are lacking? This problem is particularly important when our models are needed for conservation decision-making. For example, managers may be considering different pest-control programmes, which have the potential to lead to negative outcomes for native species, and predictions of these outcomes are needed for sound decisions to be made. These outcomes can in principle be predicted using dynamical models, however experts rarely have...Filed under: #qualitative_modelling
Pretty diagram of directory structure
 In a recent project, I wanted to create a diagram of a repository for the appendix of a paper, including comments to highlight key files and explain how the folders were organised. I found this answer on Stack Exchange by user Gonzalo Medina, which I tweaked to produce the diagram below. An annotated directory structure. The code is below: % Adapted from https://tex.stackexchange.com/a/270761 \documentclass[tikz,border=5mm]{standalone} \usepackage[edges]{forest} \definecolor{foldercolor}{RGB}{124,166,198} \newcommand{\size}[2]{ {\fontsize{#1}{0}\selectfont#2}} \definecolor{blue}{RGB}{0,50,200} \newcommand{\comment}[0]{\color{blue} \rm} \tikzset{pics/folder/.style={code={ \node[inner sep=0pt, minimum size=#1](-foldericon){}; \node[folder style, inner sep=0pt, minimum width=0.3*#1, minimum height=0.6*#1, above right, xshift=0.05*#1] at (-foldericon.west){}; \node[folder style, inner sep=0pt, minimum size=#1] at (-foldericon.center){};} }, pics/folder/.default={20pt}, folder...
In a recent project, I wanted to create a diagram of a repository for the appendix of a paper, including comments to highlight key files and explain how the folders were organised. I found this answer on Stack Exchange by user Gonzalo Medina, which I tweaked to produce the diagram below. An annotated directory structure. The code is below: % Adapted from https://tex.stackexchange.com/a/270761 \documentclass[tikz,border=5mm]{standalone} \usepackage[edges]{forest} \definecolor{foldercolor}{RGB}{124,166,198} \newcommand{\size}[2]{ {\fontsize{#1}{0}\selectfont#2}} \definecolor{blue}{RGB}{0,50,200} \newcommand{\comment}[0]{\color{blue} \rm} \tikzset{pics/folder/.style={code={ \node[inner sep=0pt, minimum size=#1](-foldericon){}; \node[folder style, inner sep=0pt, minimum width=0.3*#1, minimum height=0.6*#1, above right, xshift=0.05*#1] at (-foldericon.west){}; \node[folder style, inner sep=0pt, minimum size=#1] at (-foldericon.center){};} }, pics/folder/.default={20pt}, folder...Filed under: #coding
A Boolean approach to ecosystem structural uncertainty
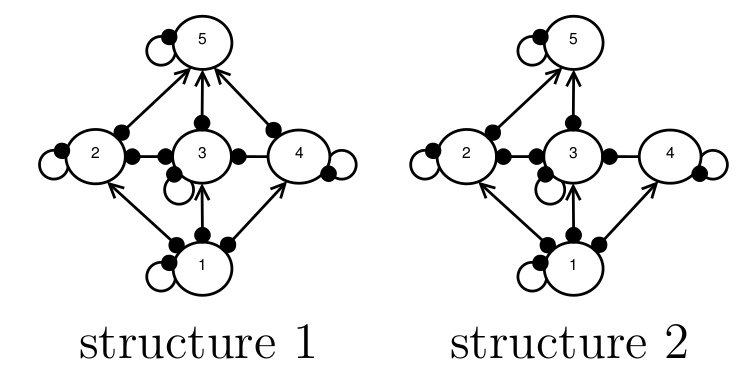 When conservation decision-makers attempt to predict the outcomes of management interventions, they frequently encounter uncertainty about the underlying ecosystem structure. Consider the example in Fig. 1, where experts are unsure whether species 4 and 5 interact. This uncertainty produces two possible structural configurations that could serve as model inputs. How might we determine which structure more accurately reflects reality? Melbourne-Thomas et al. (2012) proposed a Bayesian method to address this type of structural uncertainty. However, as I will discuss in this blog post, their approach favours simpler structures over more complex ones. Simpler structures predict fewer possible species-response outcomes. Therefore,...
When conservation decision-makers attempt to predict the outcomes of management interventions, they frequently encounter uncertainty about the underlying ecosystem structure. Consider the example in Fig. 1, where experts are unsure whether species 4 and 5 interact. This uncertainty produces two possible structural configurations that could serve as model inputs. How might we determine which structure more accurately reflects reality? Melbourne-Thomas et al. (2012) proposed a Bayesian method to address this type of structural uncertainty. However, as I will discuss in this blog post, their approach favours simpler structures over more complex ones. Simpler structures predict fewer possible species-response outcomes. Therefore,...Filed under: #qualitative_modelling
Why does it matter to conservation decision-making if alternative Qualitative Modelling methods produce contradictory predictions?
Previously, I have written about how the probabilistic approach to Qualitative Modelling (QM) (e.g. Raymond et al. 2011) can lead to contradictory predictions of species response to a management intervention, and how this is similar to the paradoxes of the Principle of Indifference that we find in the philosophy literature. A reviewer of our new manuscript (Kristensen et al. 2019) asked us an interesting and thought-provoking question: why is it that we think these contradictions matter? They did not find the contradictory predictions of the probabilistic QM methods to be a problem because it is interesting to learn how different...Filed under: #qualitative_modelling
Relationship between species discovery and extinction probability
[Update: thoughts in this post contributed to Kristensen et al. (2020).] What is the relationship between a plant’s historical probability of having being discovered and its probability that it went extinct? All else being equal, species with low abundance are less likely to be collected, and low abundance is both theoretically (e.g. McCarthy et al., 2014) and empirically cited as a good predictor of extinction probability. The empirical relationship has been observed both in general (McKinney, 1997) and specifically for plants (Sutton and Morgan, 2009; Matthies et al., 2004). It has also been found in the context of habitat fragmentation...Filed under: #undiscovered_extinctions
Off-road cycling route from Serangoon to National University of Singapore
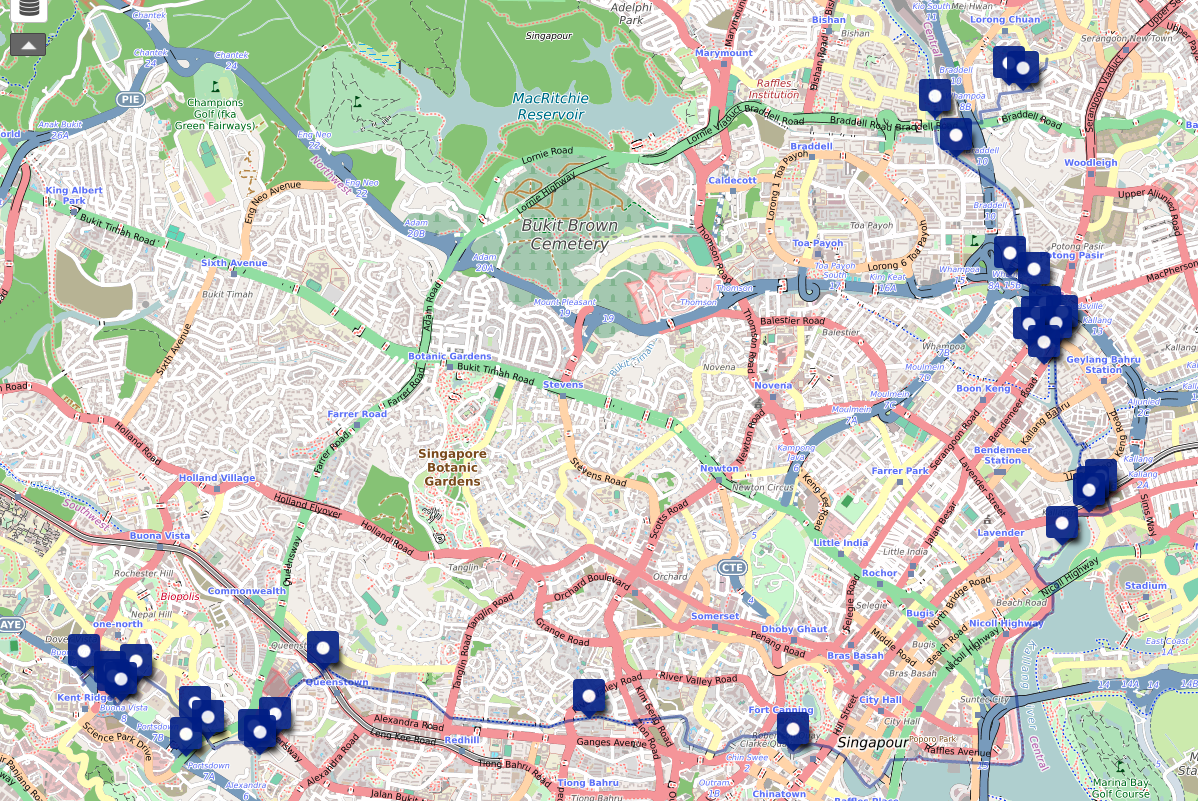 Link to annotated cycling map: NUS to Serangoon
Link to annotated cycling map: NUS to Serangoon
Filed under: #other
Guest teacher at AIS highschool
 Yesterday, I had the pleasure of teaching Year 12 students at
Australian International School about
eco-evolutionary modelling.
I created a simplified version of the mathematical model from
my paper in Proceedings of the Royal Society B
so they could solve it during class.
Yesterday, I had the pleasure of teaching Year 12 students at
Australian International School about
eco-evolutionary modelling.
I created a simplified version of the mathematical model from
my paper in Proceedings of the Royal Society B
so they could solve it during class.
Filed under: #teaching
Fixation probability of birth-death process
The goal is to understand where Eq. 2 of the Supplementary section of Sigmund et al. (2010) came from. We are considering a finite population within which individuals are pursuing different game-theoretic strategies. At each timestep, a pair of individuals is chosen at random, and they engage in a social learning process, where individual \(i\) will adopt the strategy of individual \(j\) according to a function that increases with difference in the payoff betweeen the two strategies \(P_j - P_i\), \[\frac{1}{1+\exp( -s (P_j-P_i) )}\] where \(0 \leq s < \infty\) is the imitation strength parameter. Let’s say that we have...Filed under: #cooperation
Two new papers about blue tits on Corsica
I recently read two new papers about blue tits in Corsica: Dubuc-Messier et al. (2017 Behav. Ecol.), and Dubuc-Messier et al. (2018 Evol. Biol.). The 2018 paper was interested in whether the differences between the evergreen and deciduous ecotypes on Corsica were genetic or a plastic response to the different habitat types. They took 7-12 day old nestlings and raised them in a common garden, and they found that evergreen blue tits had slower exploration speed, lower handling aggression, faster heart rate, lower body mass, and shorter tarsus. These differences were consistent with the comparison of wild adults in the...Filed under: #evolutionary_ecology
Carryover effects and local adaptation
 What allows a population in a heterogeneous landscape to become locally adapted? In general, adaptation to a rare habitat type is difficult because divergent selection is counter-acted by the homogenising effects of gene-flow. However adaptation to a rare habitat type may occur if it has a higher quality, so that a greater number of offspring can be produced there, to compensate for its relative rarity in the landscape. In a new paper published in Ecology Letters, we focused on an alternative way in which a habitat may be considered to have higher quality: by increasing the quality, rather than the...
What allows a population in a heterogeneous landscape to become locally adapted? In general, adaptation to a rare habitat type is difficult because divergent selection is counter-acted by the homogenising effects of gene-flow. However adaptation to a rare habitat type may occur if it has a higher quality, so that a greater number of offspring can be produced there, to compensate for its relative rarity in the landscape. In a new paper published in Ecology Letters, we focused on an alternative way in which a habitat may be considered to have higher quality: by increasing the quality, rather than the...Filed under: #evolutionary_ecology #dispersal
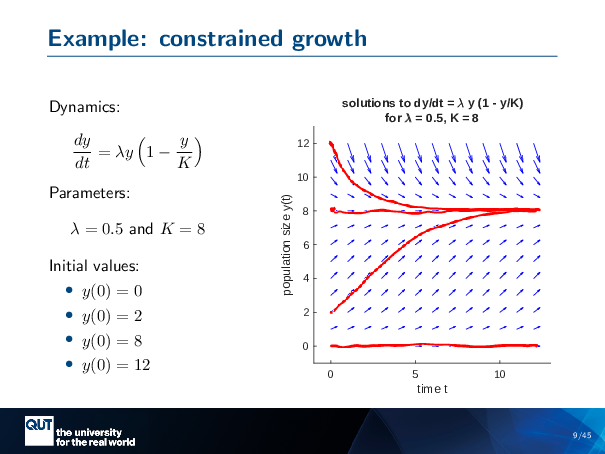
Lecture notes for LSM4255 Game Theory and Adaptive Dynamics
 These are the lecture slides for LSM4255 Game Theory and Adaptive Dynamics.
This is the short version with the point-by-point transitions removed.
Downloads:
2025: game_theory_ad4_shorter.pdf
2023: game_theory_ad2_upload.pdf
2017: game_theory_ad_upload.pdf
Preview of lecture notes.
These are the lecture slides for LSM4255 Game Theory and Adaptive Dynamics.
This is the short version with the point-by-point transitions removed.
Downloads:
2025: game_theory_ad4_shorter.pdf
2023: game_theory_ad2_upload.pdf
2017: game_theory_ad_upload.pdf
Preview of lecture notes.
Filed under: #teaching
Moments for a bivariate beta distribution
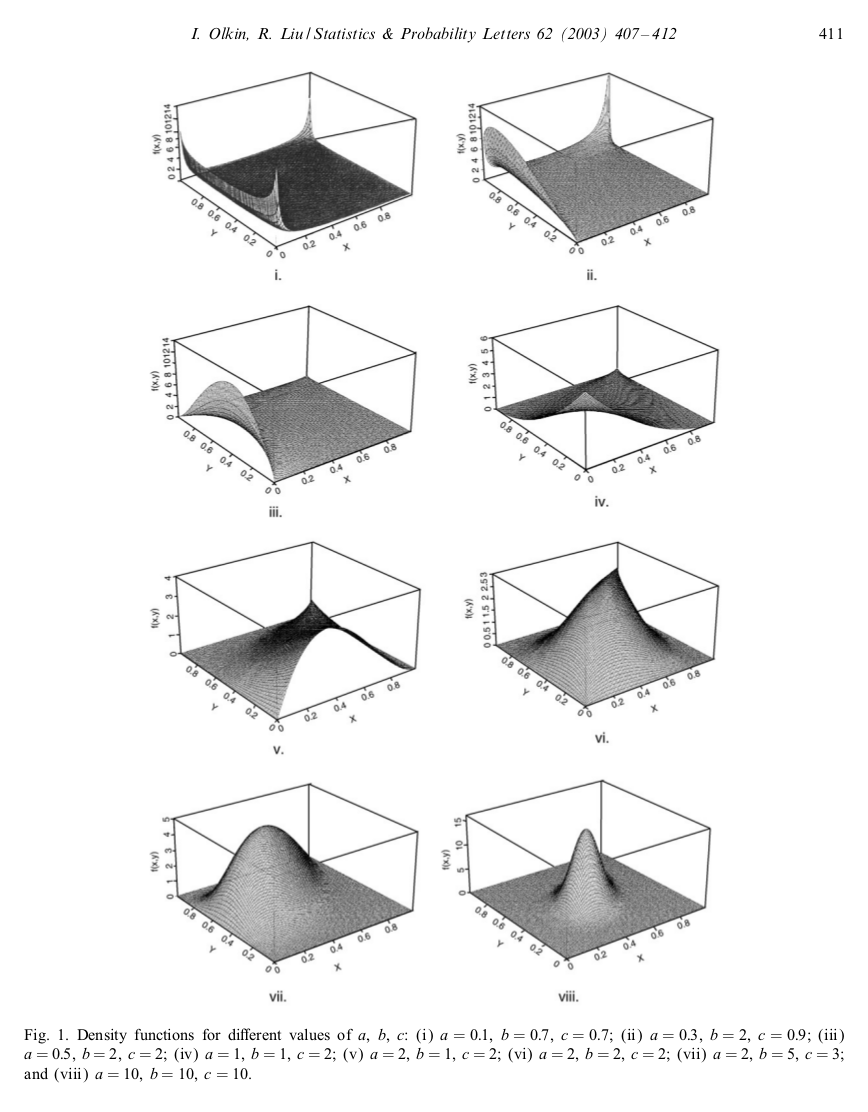 A common choice for a probability distribution of a probability is the beta distribution. It has the required support between 0 and 1, and with its two parameters we can obtain a pretty wide qualitative range for the probability density function. What should we do if we want to create correlated probabilities? We might look for some kind of multivariate generalisation of the beta distribution, one that can describe pretty flexible correlation between the variables. The generalised Dirichlet distribution allows us to describe correlated probabilities, however it has some restrictions on the support of the joint probability density function that...
A common choice for a probability distribution of a probability is the beta distribution. It has the required support between 0 and 1, and with its two parameters we can obtain a pretty wide qualitative range for the probability density function. What should we do if we want to create correlated probabilities? We might look for some kind of multivariate generalisation of the beta distribution, one that can describe pretty flexible correlation between the variables. The generalised Dirichlet distribution allows us to describe correlated probabilities, however it has some restrictions on the support of the joint probability density function that...Filed under: #undiscovered_extinctions
Comic about PI
 SMBC has a funny comic that’s also about the Principle of Indifference.
SMBC has a funny comic that’s also about the Principle of Indifference.
Filed under: #qualitative_modelling
Ecology of information
 Collaborators: Kenneth Schmidt, Jacob Johansson, François Massol, Niclas Jonzen. Breeding birds subject to nest-predation will attempt to choose a high quality nesting site with a low density of predators. For example, when recordings of chipmunk calls are played in the forest, ovenbirds will nest away from where the recordings are playing (Eureka Alert). Often models assume perfect information, so that sites are filled from highest to lowest quality, or in this case, from lowest predator density to highest density. However in reality, information about a potential nesting site must be determined by ‘prospecting’: a period of observation during which birds...
Collaborators: Kenneth Schmidt, Jacob Johansson, François Massol, Niclas Jonzen. Breeding birds subject to nest-predation will attempt to choose a high quality nesting site with a low density of predators. For example, when recordings of chipmunk calls are played in the forest, ovenbirds will nest away from where the recordings are playing (Eureka Alert). Often models assume perfect information, so that sites are filled from highest to lowest quality, or in this case, from lowest predator density to highest density. However in reality, information about a potential nesting site must be determined by ‘prospecting’: a period of observation during which birds...Filed under: #evolutionary_ecology
Modelling small insect dispersal
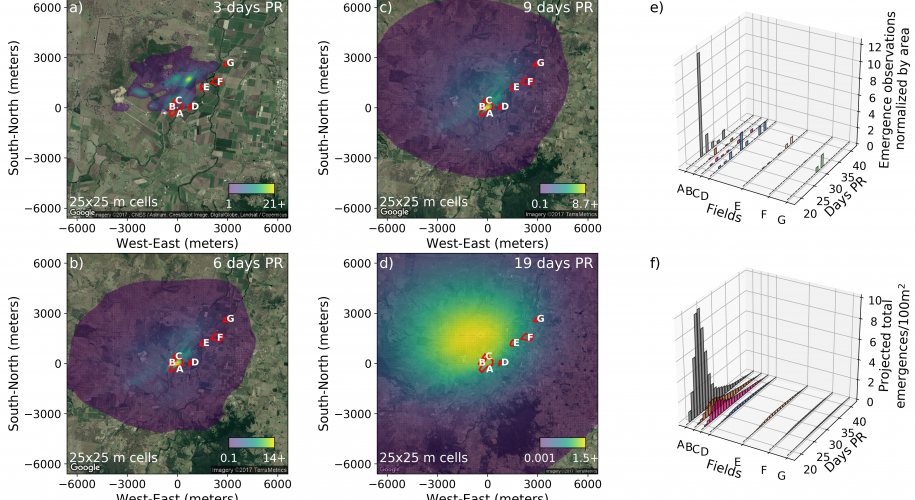 Background to the work Bemisia tabaci biotype B, commonly known as silverleaf whitefly, is a sap feeding insect that has spread globally via trade in ornamental plants. It was first detected in Australia in 1994 (Gunning et al. 1995) and has since become a crop pest and an economic problem. Eretmocerus hayati is a small parasitoid wasp that parasitises Bemisia tabaci by laying a single egg under the juveniles. Eretmocerus species have proved effective at controlling silverleaf whitefly in other countries (Goolsby et al. 2005), and so in 2004 the decision was made to release Eretmocerus hayati in Australia as...
Background to the work Bemisia tabaci biotype B, commonly known as silverleaf whitefly, is a sap feeding insect that has spread globally via trade in ornamental plants. It was first detected in Australia in 1994 (Gunning et al. 1995) and has since become a crop pest and an economic problem. Eretmocerus hayati is a small parasitoid wasp that parasitises Bemisia tabaci by laying a single egg under the juveniles. Eretmocerus species have proved effective at controlling silverleaf whitefly in other countries (Goolsby et al. 2005), and so in 2004 the decision was made to release Eretmocerus hayati in Australia as...Filed under: #dispersal
Comparing E/MSY and Chisholm method
[Update: thoughts in this post contributed to Kristensen et al. (2020).] Historical data (e.g. sighting records) can be used to estimate historical extinction rates in a variety of ways. The Chisholm et al. (2016) method (earlier post) uses the data to estimate yearly extinction probabilities. The extinctions per million species-years (E/MSY) approach (Pimm et al., 2014) estimates they extinction probability averaged over species-years. Below, I use two small examples to illustrate the similarities and differences between them. Chisholm \(=\) E/SY when extinction constant Consider the example in the table below, where the extinction probability \(\mu_t\) is the same in every...Filed under: #undiscovered_extinctions
The method of confidence belts illustrated
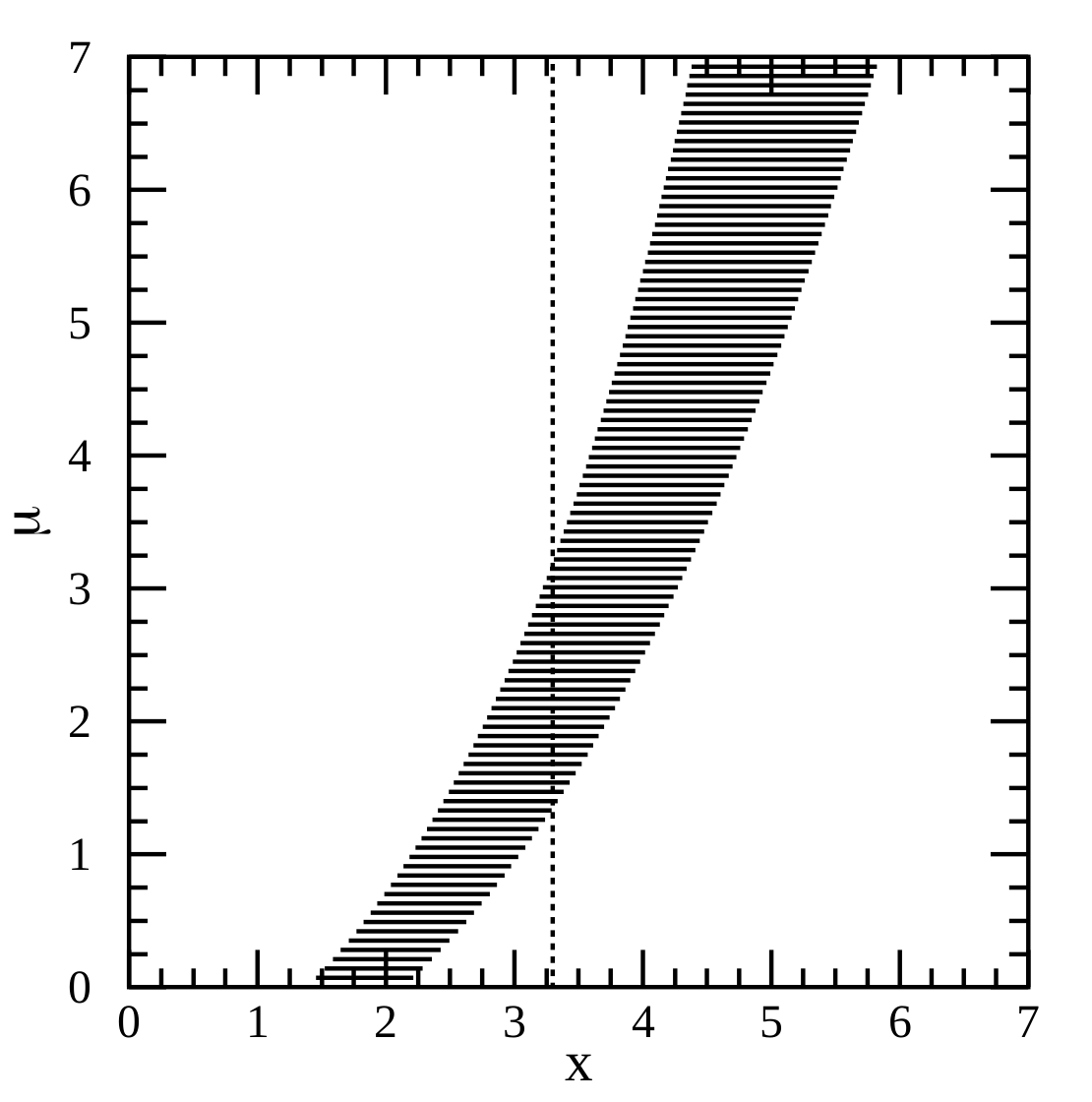 What is a confidence interval, really? We all learnt in undergrad how to find CIs for a standard distribution, but plugging numbers into equations never gave me a deep intuition for what was really going on. A worded definition is probably more helpful. Paraphrasing a bit from Wikipedia, we can think of the meaning of the confidence interval in terms of the procedure that we used to construct it. If this procedure for constructing confidence intervals is repeated on many samples, then the fraction of times that the true parameter value will fall within the different intervals will approach \(\gamma\)....
What is a confidence interval, really? We all learnt in undergrad how to find CIs for a standard distribution, but plugging numbers into equations never gave me a deep intuition for what was really going on. A worded definition is probably more helpful. Paraphrasing a bit from Wikipedia, we can think of the meaning of the confidence interval in terms of the procedure that we used to construct it. If this procedure for constructing confidence intervals is repeated on many samples, then the fraction of times that the true parameter value will fall within the different intervals will approach \(\gamma\)....Filed under: #undiscovered_extinctions
Estimating undetected extinctions
[Update: thoughts in this post contributed to Kristensen et al. (2020).] The purpose of this blog post is to give a simplified account of how the Chisholm et al. (2016) method works for estimating undetected extinctions. To estimate the historical extinction rate within a taxonomic group, a naive approach would be to divide the number of species known to be extinct by the total number of species. However, this does not account for the historical process of species discovery and temporal fluctuations in the extinction rate. The approach can be improved by estimating the cumulative probability of persistence over the...Filed under: #undiscovered_extinctions
Understanding the neigbour-modulated inclusive fitness approach
The goal is to understand Equation 4.2 of Rodrigues and Kokko (2016). To understand the technique, I read through Taylor et al. (2007), and Taylor and Frank (1996), specifically examples 4, 4a and 4b. Very briefly, we begin with a matrix \(A = [w_{i,j} ]\) whose elements represent the genetic contribution of class \(j\) to class \(i\). Then for some mutant genic value \(x\), the fitness derivative is \[\frac{dW}{dx} = \sum_{i,j} v_i \frac{dw_{i,j}}{dx} u_j\] where \(\mathbf{v}\) is the left eigenvector of \(A\) and is the reproductive values, and \(\mathbf{u}\) is the right eigenvector of \(A\) and is the class frequencies....Filed under: #cooperation
Evolution of parochialism
Previously I was reading about identity in Singapore, and I found that the government project of creating a national identity is focused upon creating a cosmopolitan mindset (i.e. one of openness and tolerance of difference), and that this interacts in different ways with the two main groups of foreigners, the ‘foreign talent’ and the ‘foreign worker’. The literature review that I did there was focused upon the sociological literature and Singapore specifically. In this post I provide a summary of some reading that I have done on experiments investigating group identity using economic games. I’ve focused specifically upon parochialism in...Filed under: #cooperation
Identity in Singapore, the Internet, and connections to game theory
The following is a literature summary that I undertook as part of my broader interest in modelling social systems, using identity in Singapore as a case study. I should preface all of this by saying that the ideas below are not ‘my own’ as such, but rather my attempt to synthesise the literature and opinions of experts in Singapore. Identity in Singapore Since independence, the Singaporean government has actively worked to construct a civic national identity for the country. Three main phases can be identified (Ortmann, 2009): (1) 1965-1980s, focused primarily upon the objective of economic growth, the idea of...Filed under: #cooperation
Termite eusociality
Termites are eusocial by possessing two subfertile or sterile castes, the worker and the soldier. The consensus is that eusociality in termites is the result of a suite of factors (Thorne, 1997), though the relative importance accorded to each in the literature has shifted over time (Howard and Thorne, 2011). For the purpose of a quick review, I have not looked into mechanisms that appear to have been set-aside in the literature, such as asymmetric relatedness and cycles of inbreeding and outbreeding (cited in Howard and Thorne, 2011). Soldiers Soldiers are monophyletic for all extant termite taxa (citations in Howard...Filed under: #cooperation
Network structural uncertainty in Qualitative Modelling
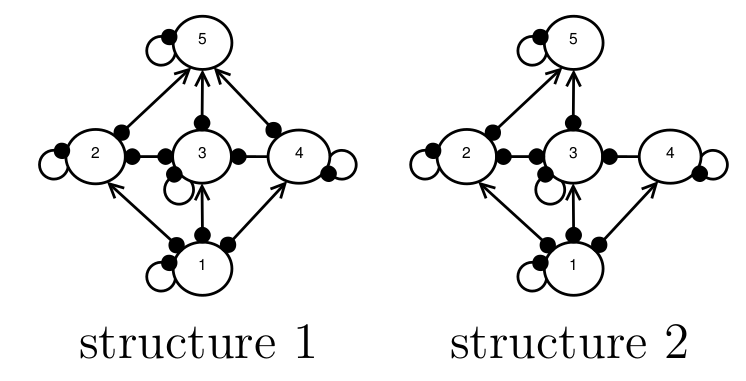 Background Imagine a situation in which we want to model the behaviour of a food web, but we don’t know what the species interaction strengths are, and we’re not 100 percent sure what the structure of the food web is, either. For example, in the figure below, experts are are uncertain about whether or not there is a direct interaction between species 4 and 5, and so both structures 1 and 2 are possibilities. We want to know, which of these candidate structures is more likely to represent the true network structure? An example of network structural uncertainty. Experts are...
Background Imagine a situation in which we want to model the behaviour of a food web, but we don’t know what the species interaction strengths are, and we’re not 100 percent sure what the structure of the food web is, either. For example, in the figure below, experts are are uncertain about whether or not there is a direct interaction between species 4 and 5, and so both structures 1 and 2 are possibilities. We want to know, which of these candidate structures is more likely to represent the true network structure? An example of network structural uncertainty. Experts are...Filed under: #qualitative_modelling
Some notes on the Principle of Indifference
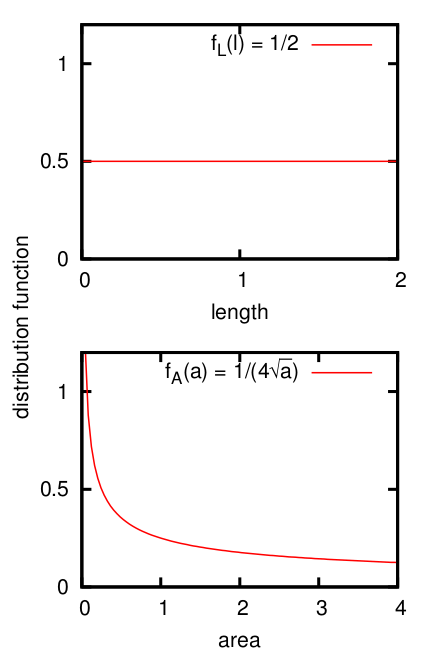 A classical statement of the Principle of Indifference (PI) is as follows (p. 45 Keynes, 1921): if there is no known reason for predicating of our subject one rather than another of several alternatives, then relatively to such knowledge the assertions of each of these alternatives have an equal probability. Thus equal probabilities must be assigned to each of several arguments, if there is an absence of positive ground for assigning unequal ones. The various forms of PI encompass two ideas (Norton, 2008). First is a truism of evidence: if we have no grounds for distinguishing outcomes, then we should...
A classical statement of the Principle of Indifference (PI) is as follows (p. 45 Keynes, 1921): if there is no known reason for predicating of our subject one rather than another of several alternatives, then relatively to such knowledge the assertions of each of these alternatives have an equal probability. Thus equal probabilities must be assigned to each of several arguments, if there is an absence of positive ground for assigning unequal ones. The various forms of PI encompass two ideas (Norton, 2008). First is a truism of evidence: if we have no grounds for distinguishing outcomes, then we should...Filed under: #qualitative_modelling
Fibonacci numbers and alternating signs in species responses to press perturbation in a food chain
In a paper from 2001, Dambacher and Rossignol made a curious observation: Fibonacci numbers appear in the adjoint and absolute feedback matrices that result from a weighted-predictions matrix type analysis (Dambacher et al. 2003) on food chains. The weighted-predictions matrix analysis is a way of predicting how species in a food web will respond to a the press perturbation of one of the species, so that implies that the pattern of species response to certain kinds of disturbance follows a neat mathematical pattern. To understand where these Fibonacci numbers come from, a paper by Usmani (1994) is useful. First some...Filed under: #food_webs #qualitative_modelling
The Principle of Indifference is actually two principles in one
 In a previous post, I wrote about the philosophical problems caused by the Principle of Indifference. The problems are illustrated with a variety of thought-experiments that create paradoxes, such as Bertrand’s paradox. I also discussed how a problem in ecological modelling for conservation decision-making seems closely related to this philosophical problem. When I realised this connection, it seemed to me that, in order to solve the ecological problem, I would need to solve the philosophical problem first. Reading the philosophy literature, however, I realised that solving the philosophical problem would not be so simple. As Shackel (2007) sums it up,...
In a previous post, I wrote about the philosophical problems caused by the Principle of Indifference. The problems are illustrated with a variety of thought-experiments that create paradoxes, such as Bertrand’s paradox. I also discussed how a problem in ecological modelling for conservation decision-making seems closely related to this philosophical problem. When I realised this connection, it seemed to me that, in order to solve the ecological problem, I would need to solve the philosophical problem first. Reading the philosophy literature, however, I realised that solving the philosophical problem would not be so simple. As Shackel (2007) sums it up,...Filed under: #qualitative_modelling
Bad for birds, good for squirrels
Here’s a nice video I stumbled upon about Lisa Aubry’s group’s work at Utah State Uni. Climate change is having a positive effect on uinta ground squirrels, allowing them to fatten-up and attain weights higher than those recorded historically. This population is survival limited, and survival probability is higher the fatter the squirrels are, so abundance responds positively to this climate change. I can’t handle how cute those squirrels are 🙂 I wonder what will happen to the rest of the ecosystem as a result of trophic interactions and other effects of increased squirrel abundance? Maybe, if a basic description...Filed under: #climate_change #evolutionary_ecology
Import self-made Sage modules and functions into Sage script?
Update Jan 2020: E.M. Bray suggests a more elegant solution in the comments, see: https://ask.sagemath.org/question/7867/importing-sage-files/?answer=48947#post-id-48947 - Let’s say that I have created a bunch of Sage code - Sage functions etc. - that I wish to reuse by importing into various Sage scripts. In straight Python, one would put these functions into their own separate file and import them using the usual from foo import funcname as localfuncname type procedure. It is unclear to me what the proper procedure for doing the same in Sage is - one can’t use the same syntax for a .sage file, and for example...Filed under: #coding
"ValueError: expected a DNF expression" when trying espresso_exprs example from pyeda docs
I’ve recently been working on a qualitative modelling project where I am trying to uncover “truths” about the response of species in an ecosystem to control of invasive species. Long story short, I’ve been looking into various boolean minimisation techniques. I’ve been playing with Python EDA, a Python library that I think provides a front-end to the Robert Brayton and Richard Rudell espresso heuristic logic minimiser, developed at University of California, Berkeley. I was trying out the examples on the Two-level Logic Minimisation docs page, and I had no issues with the second ‘Minimise truth tables’ example. However for the...Filed under: #coding #qualitative_modelling
Migratory bird phenology
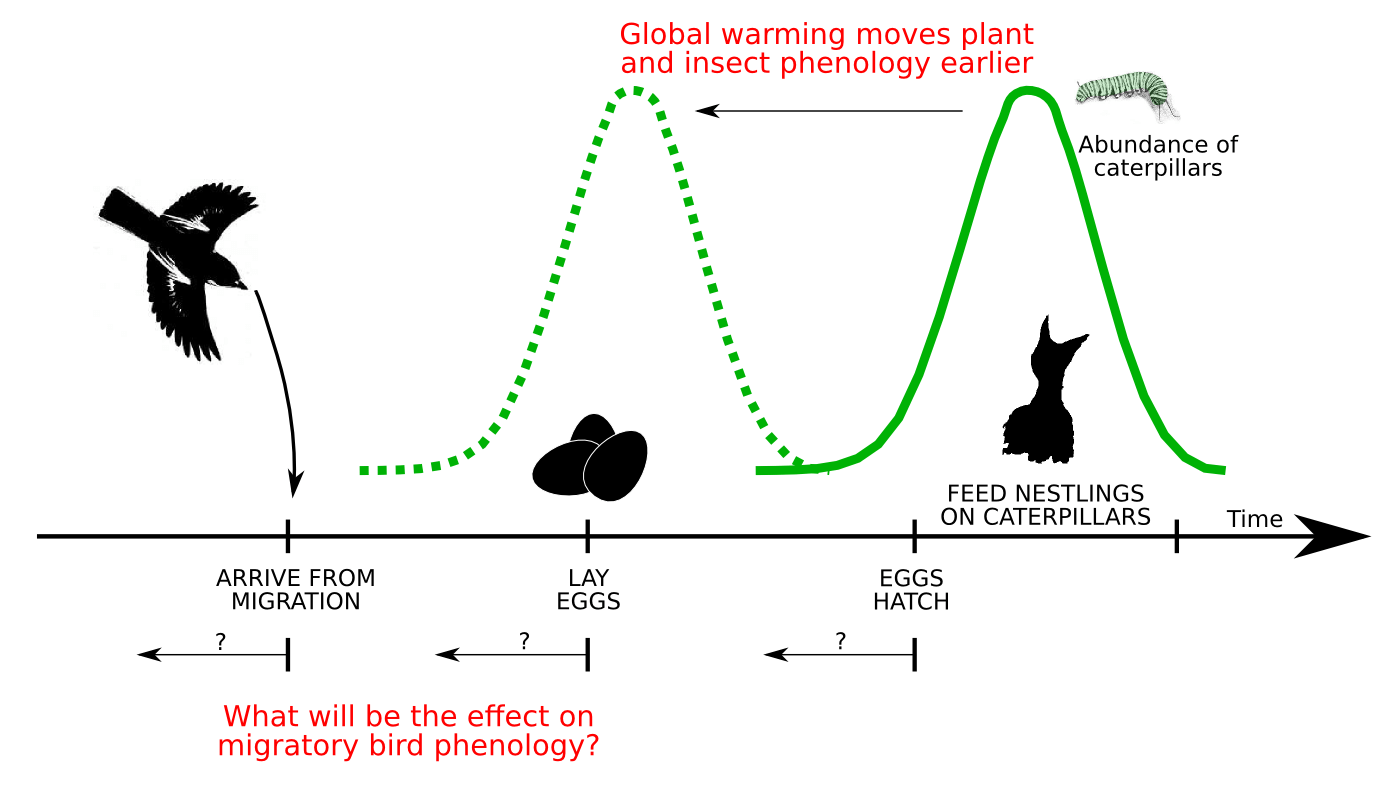 Collaborators: Jacob Johansson, Jorgen Ripa, Niclas Jonzen, Theoretical Population Ecology and Evolution Group, Lund University Kristensen, N.P., Johansson, J., Ripa, J., Jonzen, N. (In press) Phenology of two interdependent traits in migratory birds in response to climate change, Proceedings of the Royal Society B Climate change leads to an earlier arrival of spring warming. This affects the phenology of many species (Forchhammer et al. 1998, Chmielewski & Rotzer 2001, Parmesan & Yohe 2003, Edwards & Richardson 2004, Menzel et al. 2006, Beebee 2009). For example, in migratory birds, the effects can flow up the trophic levels. Earlier warm temperatures leads...
Collaborators: Jacob Johansson, Jorgen Ripa, Niclas Jonzen, Theoretical Population Ecology and Evolution Group, Lund University Kristensen, N.P., Johansson, J., Ripa, J., Jonzen, N. (In press) Phenology of two interdependent traits in migratory birds in response to climate change, Proceedings of the Royal Society B Climate change leads to an earlier arrival of spring warming. This affects the phenology of many species (Forchhammer et al. 1998, Chmielewski & Rotzer 2001, Parmesan & Yohe 2003, Edwards & Richardson 2004, Menzel et al. 2006, Beebee 2009). For example, in migratory birds, the effects can flow up the trophic levels. Earlier warm temperatures leads...Filed under: #evolutionary_ecology
The Principle of Indifference in ecological modelling
(Update April 2019: a paper on the topic below has now been published in MEE) Qualitative modelling Qualitative modelling (QM) holds the promise of obtaining predictions from dynamical models even when we don’t have all the data needed to parameterise them. How does QM achieve this? In short, the idea is to explore the range of possible parameter values to create an ensemble of possible predictions, and then interpret those predictions probabilistically and/or qualitatively. For example, if 89% of models predict that rabbit control will increase penguin populations on Macquarie Island, then that might be considered moderate support for that...Filed under: #qualitative_modelling
Lightning talk on migratory bird phenology
A lightning talk for our latest paper: Kristensen, Nadiah P., Jacob Johansson, Jörgen Ripa, and Niclas Jonzén. 2015. “Phenology of two interdependent traits in migratory birds in response to climate change.” Proceedings of the Royal Society BFiled under: #evolutionary_ecology
Biodiversity versus ecosystem services
This is a brief summary of a debate about the proper role of conservation, and the relationship between biodiversity conservation (BD) and managing for ecosystem services (ES). The debate centres around an article by Kareiva and Marvier (2012). Some threads of the discussion can be found in: A reply by Soule (2013), followed by Marvier (2014), and those below. A reply by Miller et al. (2014), followed by Kareiva (2014), Soule (2014). A reply by Doak et al. (2014b), followed by Marvier and Kareiva (2014a), Doak et al. (2014a). A reply by Cafaro and Primack (2014), followed by Marvier and...Filed under: #other
A reminder for later
The Automatic Differentiation package in Haskell can do some interesting things. Prelude Numeric.AD> jacobian ([x, y] -> [x*x*y, 5*x + sin y]) [1, 1] [[2.0,1.0],[5.0,0.5403023058681398]]Filed under: #coding
On phenology and bet-hedging
Environmental fluctuations may cause natural selection to favor phenologies that systematically deviate from the resource maximizing strategy. In temporally variable environments, because fitness is taken from the geometric (not arithmetic) mean growth rate (Gillespie, 1974), this permits the evolution of bet-hedging strategies (Simons, 2011), which are strategies that maximises total fitness by reducing temporal variation at the cost of arithmetic mean fitness (Ripa et al. 2009). Bet-hedging may either involve a single conservative bet-hedging strategy (Philippi and Seger, 1989), or where polyphenism (variable phenotypic expression of the same genotype) is possible (Moran, 1992), lead to diversified bet-hedging strategies in partially...Filed under: #evolutionary_ecology
The interesting case of blue tits in France
[Update: Four years after this blog post, I published a paper in Ecology Letters that was partly inspired by this system: Carryover effects from natal habitat type upon competitive ability lead to trait divergence or source–sink dynamics] I’ve recently been reading about a several decade long study centred around blue tits in France and on the island of Corsica. The work is being carried out by a group in the evolutionary ecology unit at the Centre D’Ecologie Fonctionnelle & Evolutive and it crosses over all kinds of interesting areas of ecology and evolution. The mainland study area is dominated by...Filed under: #evolutionary_ecology
The IPCC First Assessment Report and phenological synchrony
I just found a pretty early reference to the possibility of climate change leading to asynchrony between plant and animal phenology. From Chapter 10 page 295 of Climate Change: The IPCC Scientific Assessment (1990) Of crucial importance in plant community functioning is the synchronous operation of the life cycles of interacting plants, animals and soil organisms. Complex synchronies are found in communities in which the life cycles of plants and pollinating and seed-dispersing animals must be closely linked. Changes in climate could disrupt these synchronies. Unfortunately it didn’t attribute the idea to a reference. I wonder who was the first...Filed under: #climate_change
Are caterpillars really that important?
Typically the the kinds of models that I’m interested in assume that fledging rate is very dependent upon a phenological match between nestlings’ peak food requirements and the peak in caterpillar abundance. However I recently read a study by Cholewa and Wesołowski (2011; Acta Ornithologica) pointing out that, while literature shows that blue tits and marsh tits are indeed strongly dependent upon this food type, other tit species are more versatile and can successfully reproduce without relying upon it. Perhaps more troubling, the authors note that at the time of their writing, there was not a single study available that...Filed under: #evolutionary_ecology #climate_change
Proceedings of the Hivemind B
My partner just sent me a link to a post by Marcio von Muhlen called We Need a Github of Science. I started using Github just recently. For those new to Github (like me), it is a web-based hosting service for software development projects that use the Git revision control system. Basically you can upload your code there, and anyone else can see it and its entire revision history, comment on it, and download it. Most of interesting of all, anyone can make a copy of it to correct mistakes or to create their own extensions (forking), and you can...Filed under: #other
Add period to end of last author (Biology Letters .bst)
I had an issue today where I wanted to create a custom .bst file for Biology Letters that would format as follows: Parmesan C, Yohe G. 2003 A globally coherent fingerprint of climate change impacts across natural systems. Nature 421, 37–42. I was using makebst, incrementally editing my .dbj file, but I couldn’t seem to get that ‘.’ on the end of the author list to come out. The problem seemed to be some interaction between the default for blocks and authors. My options for author were %<>PUNCTUATION AFTER AUTHORS: So going with the default invoked (I think) this section of...Filed under: #coding
Do birds sometimes respond to warming temperatures by delaying their phenology?
[Update: thoughts in this post contributed to Kristensen et al. (2015), which found mathematical conditions under which phenology may delay.] Climate change has caused an advance in phenological events in many species. In migratory birds, the effects of warming flow causally up the trophic levels. For example, warmer temperatures lead to earlier plant phenology (e.g. budding), which leads to earlier peaks in the abundance of foods (e.g. insect larva) that are important to raising nestlings, which puts pressure upon birds to advance their own breeding timetable. In general, birds have responded to warming weather by advancing their own phenology. Migratory...Filed under: #climate_change #evolutionary_ecology
What is the relationship between lay date, mismatch, and overall fitness for migratory birds?
[Update: thoughts in this post contributed to Kristensen et al. (2015)] Food availability at the breeding site often has a peaked temporal profile such that food is very abundant for a short period of time. Therefore, migratory birds must also time their arrival such that there is adequate time to gather the resources needed for egg production, and time their nesting and laying so that nestlings can take advantage of these food peaks. Failure to do so leads to a `mismatch’, and presumably a suboptimal nestling health and fledging rate. The benefits of early laying include match to the food...Filed under: #evolutionary_ecology
The problem of arrival time and prelaying period in migratory birds
[Update: thoughts in this post contributed to Kristensen et al. (2015).] Climate change has caused an advance in phenological events in many species (Forchhammer et al. 1998, Chmielewski & Rotzer 2001, Parmesan & Yohe 2003, Edwards & Richardson 2004, Menzel et al. 2006, Beebee 2009). In migratory birds, the effects of warming flow causally up the trophic levels. For example, warmer temperatures lead to earlier plant phenology (e.g. budding) (Menzel et al. 2006, Schwartz et al. 2006, Primack et al. 2009), which leads to earlier peaks in the abundance of foods (e.g. insect larva) that are important to raising nestlings...Filed under: #evolutionary_ecology

libXmu.so.6 and libgthread-2.0.so.0 error, or getting Wolfram CDF Player working on Ubuntu
Recently a colleague sent me some information on his model and a bunch of Mathematica .nb files. I don’t use Mathematica (if I felt the need for that kind of thing I’d go for Sage) and I don’t have access to it, so I started looking around for a way to export the notebook, say to pdf. I found an online nb to pdf converter hosted by Wolfram called NBtoPDF, so I uploaded the smallest file (quite a few M) while I kept searching for a solution. I’m glad I did keep searching, because after a good deal of time...Filed under: #coding
Stability in large webs
Allesina & Tang (2012 Nature): An extension of analytic, Wigner semicircle theorem-like, stability criteria to predator-prey, competition and mutualism cases. Finds predator-prey interactions permit stability in networks as large and complex as real ones. Hierarchy is: competition-mutualism mixture, random, predator-prey. Finds that stability is less likely when both predator-prey networks and mutualistic networks are given more realistic structure (i.e. niche-model structure and nestedness respectively).Filed under: #food_webs
Large community evolution models
 Several reviews exist on the influence of evolution on food-web structure (Yoshida, 2006; Fussman et al., 2007; Loeuille & Loreau, 2009; Loeuille, 2010). In Brannstrom et al. (2008), we reviewed past theoretical modelling efforts to study food web structure, and the current state of large community evolution models to tackle the problem. Schematic illustration of the community models that were considered in our review. We started out by surveying static and dynamic ecological models, which do not possess an evolutionary component. After this, we moved on to evolution in small community models. Finally, we reviewed large community-evolution models, which synthesise...
Several reviews exist on the influence of evolution on food-web structure (Yoshida, 2006; Fussman et al., 2007; Loeuille & Loreau, 2009; Loeuille, 2010). In Brannstrom et al. (2008), we reviewed past theoretical modelling efforts to study food web structure, and the current state of large community evolution models to tackle the problem. Schematic illustration of the community models that were considered in our review. We started out by surveying static and dynamic ecological models, which do not possess an evolutionary component. After this, we moved on to evolution in small community models. Finally, we reviewed large community-evolution models, which synthesise...Filed under: #evolutionary_ecology #food_webs
Link distribution in static community models like the niche model
I was recently reading a 2006 review of food web structure by Jennifer Dunne, which includes a very thorough discussion of static community models like the niche model (Williams & Martinez 2000, Nature), and I was struck again by how unclear it is - to me, anyway - what exactly it is that these static models represent. There’s no doubt that they do an excellent job of replicating the coarse structural properties of large empirical food webs. They also provide important concrete evidence that food web structure is non-random, and a set of simple rules that replicate this structure …...Filed under: #food_webs
Basic mercurial commands
I’ve started using the Mercurial version control system. I like that it doesn’t leave a whole heap of files around, just one .hg directory. The commands to get started are fairly straightforward: hg init . hg status: Look at what is included or not. hg add *: Add everything in the directory. hg commit: Here you can enter some initial comment about the repository hg view: This allows you to look at the history of the repository through a GUIFiled under: #coding
Installing Sphinx
Many times I have found someone’s log of an installation of some new software incredibly useful, so here’s my log from installing Sphinx a while ago. First I followed the instructions in the Sphinx tutorial here, namely: Installed using sphinx-quickstart sphinx-build -b html sourcedir builddir make html This gave me a basic html page with no content I Created a python program with some rst in the docstrings, called intercept.py, which went into the source directory I added these lines to ./source/index.rst: .. automodule:: intercept :members: I also added the following lines to ./source/conf.py: * sys.path.insert(0, os.path.abspath('.')): So it could...Filed under: #coding
Pretty food web graphs
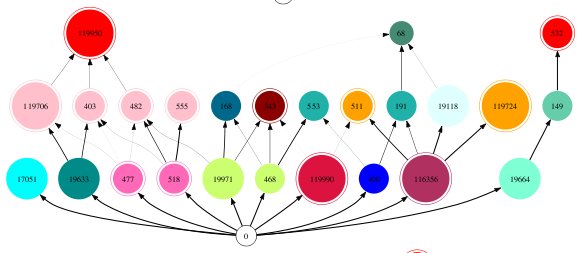 When analysing output from the Webworld food web assembly algorithm (Drossel et al. 2001, J. Theor. Biol.), I often need a good way to quickly summarise the key properties of a web. In a recent project, I created a version of Webworld in which species could invade from outside the web (randomly generated species traits, as opposed to traits derived through mutation of a native species). I wanted to know what some of my food webs looked like, including information about their lineages, so started creating figures like the one below. Webworld food web diagram generated with dotty Species are...
When analysing output from the Webworld food web assembly algorithm (Drossel et al. 2001, J. Theor. Biol.), I often need a good way to quickly summarise the key properties of a web. In a recent project, I created a version of Webworld in which species could invade from outside the web (randomly generated species traits, as opposed to traits derived through mutation of a native species). I wanted to know what some of my food webs looked like, including information about their lineages, so started creating figures like the one below. Webworld food web diagram generated with dotty Species are...Filed under: #food_webs
Fischer and the CO2 vs temperature lag
One of the most memorable moments in Al Gore’s movie “An Inconvenient Truth” occurs as Gore discusses the ice-core records for CO2 and temperature over the previous 7 “ice-ages”. After graphically showing the strong correlation between CO2 and temperature in these records, Gore is forced to mount a cherry-picker in order to physically point to the height on the graph that CO2 is predicted to reach over the next 50 years. The implication is clear: if CO2 is predicted to get so high, and given that temperature follows CO2, just imagine how high the temperature will be! In several places...Filed under: #climate_change
Webworld for Python
 A typical Webworld food web
I’ve finished up a draft module for Webworld (Drossel et al. 2001, J. Theor. Biol.) in Python that’s available for download: webworld.py
It’s also available bundled with what you’ll need for an example run: webworld.tar.gz.
A typical Webworld food web
I’ve finished up a draft module for Webworld (Drossel et al. 2001, J. Theor. Biol.) in Python that’s available for download: webworld.py
It’s also available bundled with what you’ll need for an example run: webworld.tar.gz.
Filed under: #coding #food_webs
Greenhouse gas vs temperature
 Roughly speaking, greenhouse gases have been continually increasing since 1900. They underwent a steady increase from 1900, with a sharp acceleration around 1950, followed by continued increase at this faster rate up until now. In contrast, global temperatures have oscillated. They showed a sharp increase from 1920, plateaued and then dropped around 1950, before picking up pace and increasing sharply again around 1970. Recently I was asked by someone, if greenhouse gases drive global temperatures, why haven’t temperatures shown the same pattern of increase as greenhouse gases? The work of Meehl et al. (2004, Journal of Climate), particularly the figures,...
Roughly speaking, greenhouse gases have been continually increasing since 1900. They underwent a steady increase from 1900, with a sharp acceleration around 1950, followed by continued increase at this faster rate up until now. In contrast, global temperatures have oscillated. They showed a sharp increase from 1920, plateaued and then dropped around 1950, before picking up pace and increasing sharply again around 1970. Recently I was asked by someone, if greenhouse gases drive global temperatures, why haven’t temperatures shown the same pattern of increase as greenhouse gases? The work of Meehl et al. (2004, Journal of Climate), particularly the figures,...Filed under: #climate_change
Food web assembly algorithms
 A food web assembly algorithm is a series of repeating steps by which a model food web can be created. Starting with one or a few species, new species are permitted to invade at each time step, and are lost at each time step, according to some prespecified rules. There is already a large body of food web assembly literature (Tregonning & Roberts 1979, Post & Pimm 1983, Mithen & Lawton 1986, Taylor 1988, Drake 1990, Law & Blackford 1992, Luh & Pimm 1993, Law & Morton 1996, Lockwood et al. 1997, Drossel et al. 2001, Bastolla et al. 2005,...
A food web assembly algorithm is a series of repeating steps by which a model food web can be created. Starting with one or a few species, new species are permitted to invade at each time step, and are lost at each time step, according to some prespecified rules. There is already a large body of food web assembly literature (Tregonning & Roberts 1979, Post & Pimm 1983, Mithen & Lawton 1986, Taylor 1988, Drake 1990, Law & Blackford 1992, Luh & Pimm 1993, Law & Morton 1996, Lockwood et al. 1997, Drossel et al. 2001, Bastolla et al. 2005,...Filed under: #food_webs
The change in the distance from the convex hull to the internal equilibrium during assembly
Law & Morton (unpub.) found that the distance between the interior equilibrium and the convex hull, measured as mean \(d\), increased as a permanent food web assembly progressed. I did a small set of assembly runs to test the robustness of this result, and to discover what causes \(d\) to increase. An introduction to permanence and example of how it is calculated can be found in a previous post. In verifying the result, I found that it was generally true for 2-prey, 2-predator webs regardless of the structure of the terminating web (Section 4.2). However it was not true for...Filed under: #food_webs
Permanence and the distance from the convex hull to the interior equilibrium
Background To use permanence, Lotka-Volterra dynamics have to be assumed, because it is only in this case that a sufficient condition for permanence is known: \[\dot{x}_{i} = x_i \cdot f_i(x) = x_i \cdot (r_i + (A \cdot x)_i) \quad \forall i = 1, \ldots, n.\] Such a dynamical system is permanent if two conditions hold (Hofbauer and Sigmund 1988, The theory of evolution and dynamical systems p. 98). It is dissipative; this is true for Lotka-Volterra systems where all the basal species are self-limiting and heterotrophs cannot survive without their prey. That: \(P(x) = \prod_i x_{i}^{p_i}\) for some \(p_i>0\) is...Filed under: #food_webs
An example linear programming problem in Octave
Tools for solving linear programming problems are useful to me because the necessary condition for permanence in a Lotka-Volterra system can be reduced to a linear programming problem (Jansen 1987, J. Math. Biol.; Law & Morton 1996, Ecology). Below, I’ve adapted an example from Tommi Sotinen’s ORMS 1020 lecture notes (p. 24-38) to demonstrate how to solve a linear programming problem in Octave. – Giapetto’s Woodcarving, Inc., manufactures two types of wooden toys: peace-keepers and trains. A peace-keeper sells for $27 and uses $10 worth of raw materials. Each peace-keeper that is manufactured increases Giapetto’s variable labor and overhead costs...Filed under: #coding
Portrait of a scientist
 Monica from Merici College painted a portrait of me for the 2006 Portrait of a Scientist project. It was lovely meeting Monica and it is flattering to have such a nice picture painted for me, especially one also featuring Cthulhu.
The commentary accompanying the portrait was amusing:
Monica was surprised when she met Nadiah, ‘She didn’t seem like a scientist, she was completely normal and had a nice fashion sense. She was wearing red and black - her description noted that these were her two favourite colours.
Monica from Merici College painted a portrait of me for the 2006 Portrait of a Scientist project. It was lovely meeting Monica and it is flattering to have such a nice picture painted for me, especially one also featuring Cthulhu.
The commentary accompanying the portrait was amusing:
Monica was surprised when she met Nadiah, ‘She didn’t seem like a scientist, she was completely normal and had a nice fashion sense. She was wearing red and black - her description noted that these were her two favourite colours.
Filed under: #other
Invasibility and food web structure in assembly algorithms
The following is a small literature search I did during my post-doc at CSIRO. The goal was to find system-level food web attributes that are correlated with invasibility. System-level food web attributes It is worth doing a quick overview of what is meant by the “system-level food web attributes” mentioned in the objective. Using the early empirical food web literature, a group of authors noted commonalities in certain attributes across many different webs (Cohen, 1989; Briand & Cohen, 1984,1987). Attributes included: the fraction of species in each trophic type, the fraction of links between each trophic type, link density and...Filed under: #food_webs
subscribe via RSS