Filed Under #evolutionary_ecology
Jul 1, 2022
 I recently came across a paper by Kaj Hulthen and others in Journal of Animal Ecology showing good empirical evidence for a model I coauthored a few years ago with Anna Harts and Hanna Kokko. Hulthen et al. (2022) were interested in the migration timing of roach (Rutilus rutilus). Specifically, they were interested in how the combination of selection for early arrival plus high predation risk could explain the relatively high migration synchrony in spring compared to autumn. Roach migrate from lake to stream in autumn, and from stream to lake in spring. In the lake in spring, zooplankton numbers...
I recently came across a paper by Kaj Hulthen and others in Journal of Animal Ecology showing good empirical evidence for a model I coauthored a few years ago with Anna Harts and Hanna Kokko. Hulthen et al. (2022) were interested in the migration timing of roach (Rutilus rutilus). Specifically, they were interested in how the combination of selection for early arrival plus high predation risk could explain the relatively high migration synchrony in spring compared to autumn. Roach migrate from lake to stream in autumn, and from stream to lake in spring. In the lake in spring, zooplankton numbers...
Predator dilution effect synchronises fish migration
I recently came across a paper by Kaj Hulthen and others in Journal of Animal Ecology showing good empirical evidence for a model I coauthored a few years ago with Anna Harts and Hanna Kokko. Hulthen et al. (2022) were interested in the migration timing of roach (Rutilus rutilus). Specifically, they were interested in how the combination of selection for early arrival plus high predation risk could explain the relatively high migration synchrony in spring compared to autumn. Roach migrate from lake to stream in autumn, and from stream to lake in spring. In the lake in spring, zooplankton numbers...
Aug 9, 2019
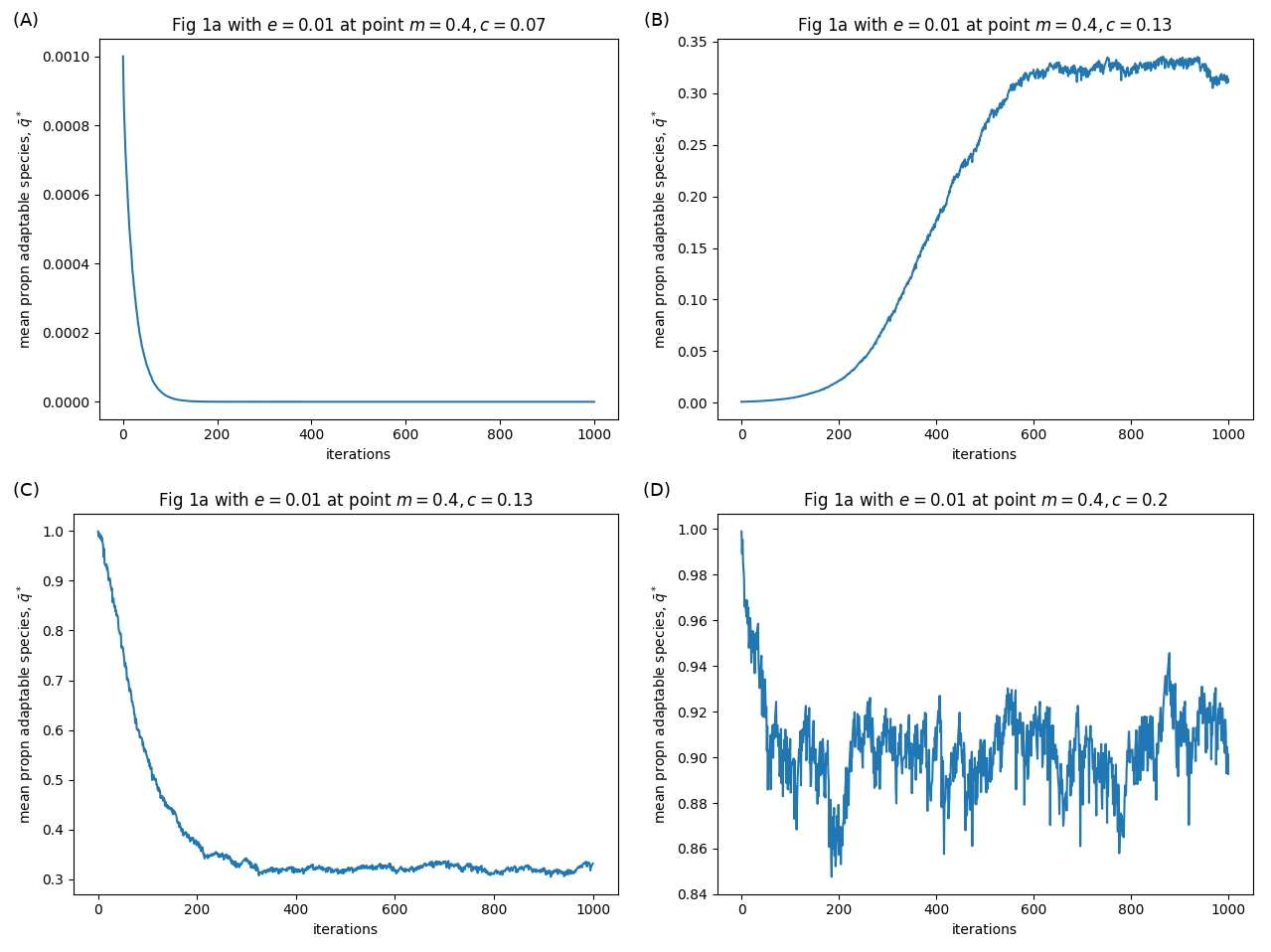 I recently read a paper by Kawecki (2017), which presents a new mechanism for something analogous to fugitive coexistence. The paper has a really great literature overview, which I won’t be able to do justice here. In short, fugitive coexistence is when an inferior species persists on a patchy landscape by being a better coloniser: when a local extinction occurs, they are quicker to arrive at the patch and so exploit the time window before the superior competitor arrives. But Kawecki had another idea about how coexistence could occur. In the paper, Kawecki explores the coexistence of a ‘jack of...
I recently read a paper by Kawecki (2017), which presents a new mechanism for something analogous to fugitive coexistence. The paper has a really great literature overview, which I won’t be able to do justice here. In short, fugitive coexistence is when an inferior species persists on a patchy landscape by being a better coloniser: when a local extinction occurs, they are quicker to arrive at the patch and so exploit the time window before the superior competitor arrives. But Kawecki had another idea about how coexistence could occur. In the paper, Kawecki explores the coexistence of a ‘jack of...
Playing with a new model for fugitive coexistence
I recently read a paper by Kawecki (2017), which presents a new mechanism for something analogous to fugitive coexistence. The paper has a really great literature overview, which I won’t be able to do justice here. In short, fugitive coexistence is when an inferior species persists on a patchy landscape by being a better coloniser: when a local extinction occurs, they are quicker to arrive at the patch and so exploit the time window before the superior competitor arrives. But Kawecki had another idea about how coexistence could occur. In the paper, Kawecki explores the coexistence of a ‘jack of...
Aug 1, 2018
Two new papers about blue tits on Corsica
I recently read two new papers about blue tits in Corsica: Dubuc-Messier et al. (2017 Behav. Ecol.), and Dubuc-Messier et al. (2018 Evol. Biol.). The 2018 paper was interested in whether the differences between the evergreen and deciduous ecotypes on Corsica were genetic or a plastic response to the different habitat types. They took 7-12 day old nestlings and raised them in a common garden, and they found that evergreen blue tits had slower exploration speed, lower handling aggression, faster heart rate, lower body mass, and shorter tarsus. These differences were consistent with the comparison of wild adults in the...Jun 26, 2018
 What allows a population in a heterogeneous landscape to become locally adapted? In general, adaptation to a rare habitat type is difficult because divergent selection is counter-acted by the homogenising effects of gene-flow. However adaptation to a rare habitat type may occur if it has a higher quality, so that a greater number of offspring can be produced there, to compensate for its relative rarity in the landscape. In a new paper published in Ecology Letters, we focused on an alternative way in which a habitat may be considered to have higher quality: by increasing the quality, rather than the...
What allows a population in a heterogeneous landscape to become locally adapted? In general, adaptation to a rare habitat type is difficult because divergent selection is counter-acted by the homogenising effects of gene-flow. However adaptation to a rare habitat type may occur if it has a higher quality, so that a greater number of offspring can be produced there, to compensate for its relative rarity in the landscape. In a new paper published in Ecology Letters, we focused on an alternative way in which a habitat may be considered to have higher quality: by increasing the quality, rather than the...
Carryover effects and local adaptation
What allows a population in a heterogeneous landscape to become locally adapted? In general, adaptation to a rare habitat type is difficult because divergent selection is counter-acted by the homogenising effects of gene-flow. However adaptation to a rare habitat type may occur if it has a higher quality, so that a greater number of offspring can be produced there, to compensate for its relative rarity in the landscape. In a new paper published in Ecology Letters, we focused on an alternative way in which a habitat may be considered to have higher quality: by increasing the quality, rather than the...
Jul 23, 2017
 Collaborators: Kenneth Schmidt, Jacob Johansson, François Massol, Niclas Jonzen. Breeding birds subject to nest-predation will attempt to choose a high quality nesting site with a low density of predators. For example, when recordings of chipmunk calls are played in the forest, ovenbirds will nest away from where the recordings are playing (Eureka Alert). Often models assume perfect information, so that sites are filled from highest to lowest quality, or in this case, from lowest predator density to highest density. However in reality, information about a potential nesting site must be determined by ‘prospecting’: a period of observation during which birds...
Collaborators: Kenneth Schmidt, Jacob Johansson, François Massol, Niclas Jonzen. Breeding birds subject to nest-predation will attempt to choose a high quality nesting site with a low density of predators. For example, when recordings of chipmunk calls are played in the forest, ovenbirds will nest away from where the recordings are playing (Eureka Alert). Often models assume perfect information, so that sites are filled from highest to lowest quality, or in this case, from lowest predator density to highest density. However in reality, information about a potential nesting site must be determined by ‘prospecting’: a period of observation during which birds...
Ecology of information
Collaborators: Kenneth Schmidt, Jacob Johansson, François Massol, Niclas Jonzen. Breeding birds subject to nest-predation will attempt to choose a high quality nesting site with a low density of predators. For example, when recordings of chipmunk calls are played in the forest, ovenbirds will nest away from where the recordings are playing (Eureka Alert). Often models assume perfect information, so that sites are filled from highest to lowest quality, or in this case, from lowest predator density to highest density. However in reality, information about a potential nesting site must be determined by ‘prospecting’: a period of observation during which birds...
Aug 18, 2015
Bad for birds, good for squirrels
Here’s a nice video I stumbled upon about Lisa Aubry’s group’s work at Utah State Uni. Climate change is having a positive effect on uinta ground squirrels, allowing them to fatten-up and attain weights higher than those recorded historically. This population is survival limited, and survival probability is higher the fatter the squirrels are, so abundance responds positively to this climate change. I can’t handle how cute those squirrels are 🙂 I wonder what will happen to the rest of the ecosystem as a result of trophic interactions and other effects of increased squirrel abundance? Maybe, if a basic description...May 22, 2015
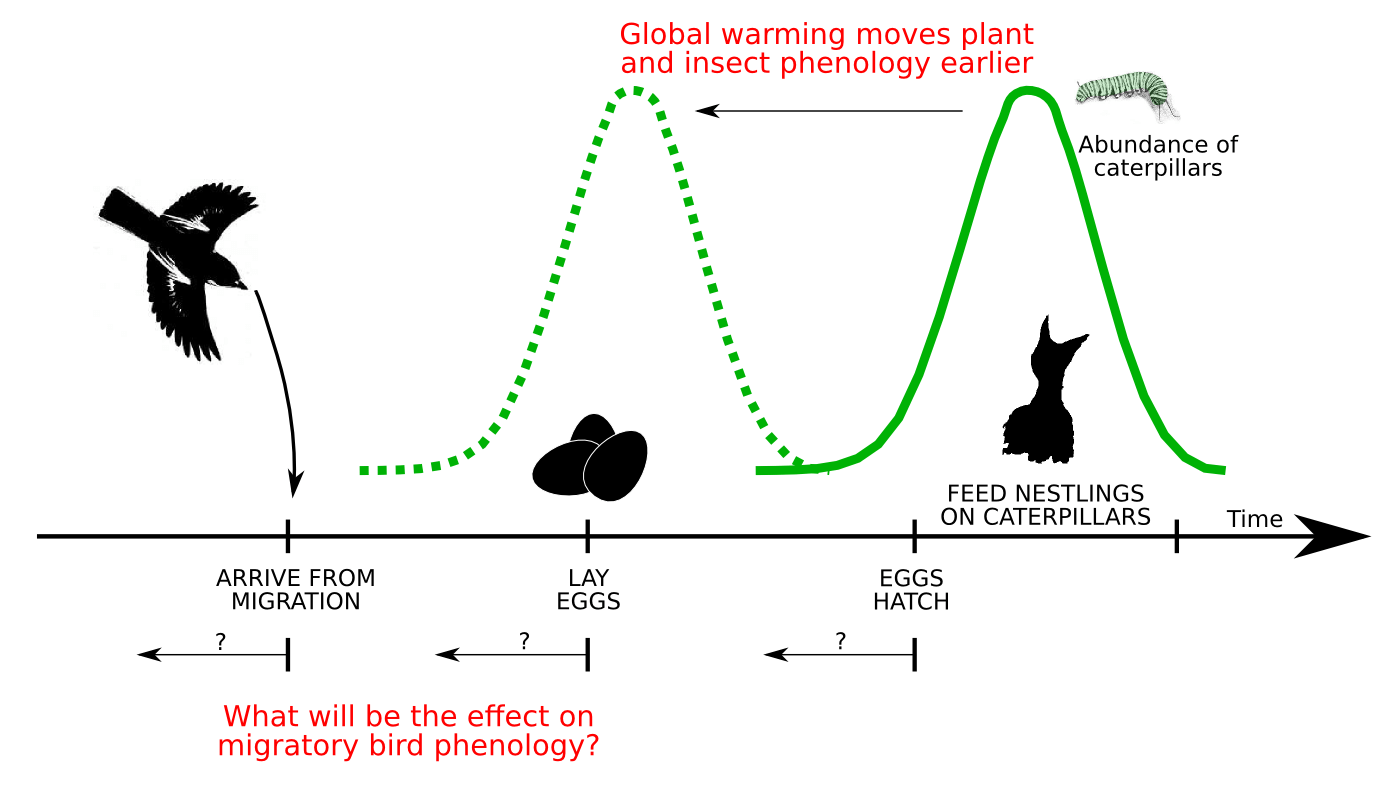 Collaborators: Jacob Johansson, Jorgen Ripa, Niclas Jonzen, Theoretical Population Ecology and Evolution Group, Lund University Kristensen, N.P., Johansson, J., Ripa, J., Jonzen, N. (In press) Phenology of two interdependent traits in migratory birds in response to climate change, Proceedings of the Royal Society B Climate change leads to an earlier arrival of spring warming. This affects the phenology of many species (Forchhammer et al. 1998, Chmielewski & Rotzer 2001, Parmesan & Yohe 2003, Edwards & Richardson 2004, Menzel et al. 2006, Beebee 2009). For example, in migratory birds, the effects can flow up the trophic levels. Earlier warm temperatures leads...
Collaborators: Jacob Johansson, Jorgen Ripa, Niclas Jonzen, Theoretical Population Ecology and Evolution Group, Lund University Kristensen, N.P., Johansson, J., Ripa, J., Jonzen, N. (In press) Phenology of two interdependent traits in migratory birds in response to climate change, Proceedings of the Royal Society B Climate change leads to an earlier arrival of spring warming. This affects the phenology of many species (Forchhammer et al. 1998, Chmielewski & Rotzer 2001, Parmesan & Yohe 2003, Edwards & Richardson 2004, Menzel et al. 2006, Beebee 2009). For example, in migratory birds, the effects can flow up the trophic levels. Earlier warm temperatures leads...
Migratory bird phenology
Collaborators: Jacob Johansson, Jorgen Ripa, Niclas Jonzen, Theoretical Population Ecology and Evolution Group, Lund University Kristensen, N.P., Johansson, J., Ripa, J., Jonzen, N. (In press) Phenology of two interdependent traits in migratory birds in response to climate change, Proceedings of the Royal Society B Climate change leads to an earlier arrival of spring warming. This affects the phenology of many species (Forchhammer et al. 1998, Chmielewski & Rotzer 2001, Parmesan & Yohe 2003, Edwards & Richardson 2004, Menzel et al. 2006, Beebee 2009). For example, in migratory birds, the effects can flow up the trophic levels. Earlier warm temperatures leads...
Apr 22, 2015
Lightning talk on migratory bird phenology
A lightning talk for our latest paper: Kristensen, Nadiah P., Jacob Johansson, Jörgen Ripa, and Niclas Jonzén. 2015. “Phenology of two interdependent traits in migratory birds in response to climate change.” Proceedings of the Royal Society BOct 21, 2014
On phenology and bet-hedging
Environmental fluctuations may cause natural selection to favor phenologies that systematically deviate from the resource maximizing strategy. In temporally variable environments, because fitness is taken from the geometric (not arithmetic) mean growth rate (Gillespie, 1974), this permits the evolution of bet-hedging strategies (Simons, 2011), which are strategies that maximises total fitness by reducing temporal variation at the cost of arithmetic mean fitness (Ripa et al. 2009). Bet-hedging may either involve a single conservative bet-hedging strategy (Philippi and Seger, 1989), or where polyphenism (variable phenotypic expression of the same genotype) is possible (Moran, 1992), lead to diversified bet-hedging strategies in partially...Jun 10, 2014
The interesting case of blue tits in France
[Update: Four years after this blog post, I published a paper in Ecology Letters that was partly inspired by this system: Carryover effects from natal habitat type upon competitive ability lead to trait divergence or source–sink dynamics] I’ve recently been reading about a several decade long study centred around blue tits in France and on the island of Corsica. The work is being carried out by a group in the evolutionary ecology unit at the Centre D’Ecologie Fonctionnelle & Evolutive and it crosses over all kinds of interesting areas of ecology and evolution. The mainland study area is dominated by...May 21, 2014
Are caterpillars really that important?
Typically the the kinds of models that I’m interested in assume that fledging rate is very dependent upon a phenological match between nestlings’ peak food requirements and the peak in caterpillar abundance. However I recently read a study by Cholewa and Wesołowski (2011; Acta Ornithologica) pointing out that, while literature shows that blue tits and marsh tits are indeed strongly dependent upon this food type, other tit species are more versatile and can successfully reproduce without relying upon it. Perhaps more troubling, the authors note that at the time of their writing, there was not a single study available that...Apr 9, 2013
Do birds sometimes respond to warming temperatures by delaying their phenology?
[Update: thoughts in this post contributed to Kristensen et al. (2015), which found mathematical conditions under which phenology may delay.] Climate change has caused an advance in phenological events in many species. In migratory birds, the effects of warming flow causally up the trophic levels. For example, warmer temperatures lead to earlier plant phenology (e.g. budding), which leads to earlier peaks in the abundance of foods (e.g. insect larva) that are important to raising nestlings, which puts pressure upon birds to advance their own breeding timetable. In general, birds have responded to warming weather by advancing their own phenology. Migratory...Jan 10, 2013
What is the relationship between lay date, mismatch, and overall fitness for migratory birds?
[Update: thoughts in this post contributed to Kristensen et al. (2015)] Food availability at the breeding site often has a peaked temporal profile such that food is very abundant for a short period of time. Therefore, migratory birds must also time their arrival such that there is adequate time to gather the resources needed for egg production, and time their nesting and laying so that nestlings can take advantage of these food peaks. Failure to do so leads to a `mismatch’, and presumably a suboptimal nestling health and fledging rate. The benefits of early laying include match to the food...Oct 13, 2012
The problem of arrival time and prelaying period in migratory birds
[Update: thoughts in this post contributed to Kristensen et al. (2015).] Climate change has caused an advance in phenological events in many species (Forchhammer et al. 1998, Chmielewski & Rotzer 2001, Parmesan & Yohe 2003, Edwards & Richardson 2004, Menzel et al. 2006, Beebee 2009). In migratory birds, the effects of warming flow causally up the trophic levels. For example, warmer temperatures lead to earlier plant phenology (e.g. budding) (Menzel et al. 2006, Schwartz et al. 2006, Primack et al. 2009), which leads to earlier peaks in the abundance of foods (e.g. insect larva) that are important to raising nestlings...Feb 16, 2012
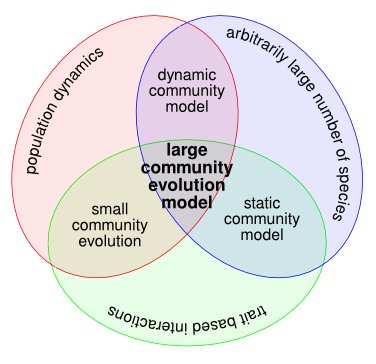 Several reviews exist on the influence of evolution on food-web structure (Yoshida, 2006; Fussman et al., 2007; Loeuille & Loreau, 2009; Loeuille, 2010). In Brannstrom et al. (2008), we reviewed past theoretical modelling efforts to study food web structure, and the current state of large community evolution models to tackle the problem. Schematic illustration of the community models that were considered in our review. We started out by surveying static and dynamic ecological models, which do not possess an evolutionary component. After this, we moved on to evolution in small community models. Finally, we reviewed large community-evolution models, which synthesise...
Several reviews exist on the influence of evolution on food-web structure (Yoshida, 2006; Fussman et al., 2007; Loeuille & Loreau, 2009; Loeuille, 2010). In Brannstrom et al. (2008), we reviewed past theoretical modelling efforts to study food web structure, and the current state of large community evolution models to tackle the problem. Schematic illustration of the community models that were considered in our review. We started out by surveying static and dynamic ecological models, which do not possess an evolutionary component. After this, we moved on to evolution in small community models. Finally, we reviewed large community-evolution models, which synthesise...
Large community evolution models
Several reviews exist on the influence of evolution on food-web structure (Yoshida, 2006; Fussman et al., 2007; Loeuille & Loreau, 2009; Loeuille, 2010). In Brannstrom et al. (2008), we reviewed past theoretical modelling efforts to study food web structure, and the current state of large community evolution models to tackle the problem. Schematic illustration of the community models that were considered in our review. We started out by surveying static and dynamic ecological models, which do not possess an evolutionary component. After this, we moved on to evolution in small community models. Finally, we reviewed large community-evolution models, which synthesise...